
Bennettitales
| Bennettitales | |
|---|---|

| |
| Restoration of a member of Williamsoniaceae by Thérèse Ekblom | |

| |
| Life restoration of "Williamsonia" sewardiana from the Early Cretaceous of India, which may represent an early member of Cycadeoidaceae | |
| Scientific classification | |
| Kingdom: | Plantae |
| Clade: | Tracheophytes |
| Clade: | Spermatophytes |
| Order: | †Bennettitales Engler, 1892 |
| Families | |
Bennettitales (also known as cycadeoids) is an extinct
Although certainly gymnosperms sensu lato (cone-bearing seed plants), the relationships of bennettitaleans to other seed plants is debated. Their general resemblance to cycads is contradicted by numerous more subtle features of their reproductive systems and leaf structure. Some authors have linked bennettitaleans to angiosperms (flowering plants) and gnetophytes (a rare and unusual group of modern gymnosperms), forming a broader group known as Anthophyta. Molecular data contradicts this, with gnetophytes found to be much more genetically similar to conifers. The exact position of Bennettitales remains uncertain.
Description


Bennettitales are divided into two families, Cycadeoidaceae and Williamsoniaceae, which have distinct growth habits. Cycadeoidaceae had stout, cycad-like trunks with bisporangiate (containing both megaspores and microspores) strobili (cones) serving as their reproductive structures. Williamsoniaceae either had bisporangiate or monosporangiate cones, and distinctly slender and branching woody trunks.[1] The Williamsoniaceae grew as woody shrubs with a divaricate branching habit, similar to that of Banksia.[2] It has been suggested that Williamsoniaceae are a paraphyletic (not containing all descendants of a common ancestor) assemblage of all Bennettitales that do not belong to the Cycadeoidaceae.[2]
Foliage
In general, bennettitalean leaves are attached to the stem with a helical (corkscrew) arrangement. Some leaves (most species of
The foliage of bennettitaleans resembles that of cycads to such an extent that the foliage of the two groups cannot be reliably distinguished based on gross morphology alone. However, fossil foliage which preserves the cuticle can be assigned to either group with confidence. The stomata of bennettitaleans are described as syndetocheilic. This means that the main paired guard cells develop from the same mother cells as the subsidiary cells which surround them. This contrasts with the haplocheilic stomata of cycads and conifers. In haplocheilic stomata, the ring of subsidiary cells are not derived from the same original structures as the guard cells. This fundamental difference is the main way to differentiate bennettitalean and cycad foliage.[5]
Cones and seeds


Like other gymnosperms, bennettitalean reproductive inflorescences come in the form of
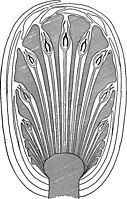
Many bennettitaleans are bisporangiate, where the pollen and ovules are hosted on the same (bisexual or hermaphrodite) cone. Cavities filled with curved synangia-bearing microsporophylls are encased by thin radiating structures, including thick, infertile interseminal scales and fertile sporophylls with ovules at their tips. The presence of ovules at the tips of sporophylls, rather than the tips of stems, is a major difference between the cones of bennettitaleans and gnetophytes. As the cone is fertilized and matures, the microsporophylls wither away and the ovules transform into seeds.[6]
Most bennettitaleans in the family Williamsoniaceae are instead monosporangiate, with separate pollen and ovule-producing (unisexual) cones on the same plant. The ovule-producing (female) cones (Williamsonia, etc.) are similar to mature bisporangiate cones, with interseminal scales and ovule-tipped sporophylls enclosed by bracts. Pollen-producing (male) cones (Weltrichia, etc.), on the other hand, feature an exposed crown of tapering microsporophylls with adaxial rows of synangia. The microsporophylls may host a single linear row of paired synangia, or instead synangia arranged in a pinnate (feather-shaped) pattern.[6]
Cycadeoidaceans have been suggested to have been self-pollinating, with their stems and cones buried underground,
Several groups of Jurassic and Early Cretaceous insects possessed a long proboscis, and it has been suggested that they fed on nectar produced by bennettitalean reproductive structures, such as the bisexual williamsoniacean reproductive structure Williamsoniella, which had a long, narrow central receptacle which was likely otherwise inaccessible.[10] Early Cretaceous bennettitalean pollen has been found directly associated with a proboscis bearing fly belonging to the extinct family Zhangsolvidae, providing evidence that this family acted as pollinators for the group.[11] The interseminal scales of Bennettitales ovulate cones may have become fleshy at maturity, which could have potentially made then attractive to wild animals that served as seed dispersers.[12]
Taxonomy
History of discovery
The Cycadeoideaceae (originally “Cycadeoideae”) were named by English geologist
Relationships to other seed plants
The
Evolutionary history
The oldest confirmed fossils of bennettitaleans are leaves of Nilssoniopteris shanxiensis, a species from the upper part of the Upper Shihhotse Formation in Shanxi Province, China.[5] This strata is dated to the early Kungurian stage of the early Permian (Cisuralian), around 281 million years ago.[26] Supposed Carboniferous-Permian records of Pterophyllum do not have conclusive bennettitalean affinities or have been reinterpreted as cycad foliage in the form genus Pseudoctenis.[27] True Permian records of benettitalean leaves are rare; outside of the Shihhotse Formation they are only found in the Late Permian (likely Changhsingian)-age Umm Irna Formation in Jordan.[5] This formation is notable for the early occurrence of other Mesozoic-style flora, including the earliest records of corystospermalean foliage (Dicroidium).[28] The order Fredlindiales (containing the genus Fredlindia) from the Late Triassic of Gondwana appears to be closely related to Bennettitales, but differs from it in some aspects of its reproductive organs.[16]
The bennettitalean fossil record reappeared in the Middle Triassic, and williamsoniaceans became globally distributed by the end of the period.[29][5] The oldest bennettitalean reproductive structures are small Williamsonia "flowers" from the Middle Triassic Esk Formation of Australia.[16] While Williamsoniaceae had a global distribution, Cycadeoidaceae appear to have been primarily confined to the western parts of Laurasia, and are primarily known from the Cretaceous.[16] Bennettitales were widespread and abundant during the Jurassic and Early Cretaceous, however Bennettitales severely declined during the Late Cretaceous, coincident with the rise of flowering plants, being mostly extinct by the end of the period, with the final known remains from the Northern Hemisphere being found in the polar latitude Kakanaut Formation in Chukotka, Russia, dating to the Maastrichtian, assignable to Pterophyllum.[30] A possible late record has been reported from the early Oligocene of eastern Australia and Tasmania, assignable to the genus Ptilophyllum, but no cuticle was preserved, making the referral inconclusive.[31]
Subgroups
- Cycadolepis (scales or bracts, unplaced in family)
- Haitingeria? (pollen organ?)
- Lunzia (pollen organ, unplaced in family)
- Leguminanthus? (pollen organ?)
- Leuthardtia? (pollen organ?)
- Westersheimia (ovulate organ, unplaced in family)
- Anthrophyopsis?[32] (leaf)
- Family Cycadeoidaceae
- Family Williamsoniaceae
- Anomozamites (leaf)
- Bennetticarpus (female seed cone)
- Bennettistemon (male pollen organ)
- Bucklandia (axes)
- Eoginkgoites (leaf)
- Ischnophyton
- Kimuriella (whole plant)
- Nilssoniopteris (leaf)
- Otozamites (leaf)
- Pterophyllum (leaf)
- Ptilophyllum (leaf)
- Vardekloeftia (female seed cone)
- Weltrichia (male pollen organ)
- Wielandiella (whole plant)
- Williamsonia (female seed cones)
- Williamsoniella (bisexual reproductive structure)
- Zamites (leaf, in partim)
- Haitingeria (pollen organ)
Bennettitales is typically considered the sole order in the class Bennettitopsida Engler (1897) or Cycadeoideopsida Scott (1923). Most paleobotanists prefer the two families as used here, though some authors, such as Anderson & Anderson (2007), classify the order via a larger number of families.[33] Anderson & Anderson also classified the orders Fredlindiales Anderson & Anderson (2003)[34] and Pentoxylales Pilger & Melchior (1954) within Bennettitopsida.[33]
Gallery
-
 Fossil leaf of Zamites mariposana from the Jurassic.
Fossil leaf of Zamites mariposana from the Jurassic. -
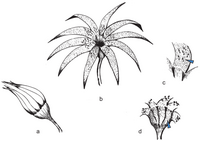 Restoration of Weltrichia magna from the Jurassic of Mexico
Restoration of Weltrichia magna from the Jurassic of Mexico -
 Otozamites leaf
Otozamites leaf -
 Leaf of Anomozamites
Leaf of Anomozamites -
 Seed bearingBennettites
Seed bearingBennettites -
 Trunk of Cycadeoidea
Trunk of Cycadeoidea -
 Fossil of Ptilophyllum grandifolium
Fossil of Ptilophyllum grandifolium




_(20595091038).jpg)


References
- ^ S2CID 13022302.
- ^ S2CID 84440045.
- S2CID 232349931.
- S2CID 44559347.
- ^ .
- ^ PMID 21628190.
- ISBN 978-0-253-00199-3.
- ^ Frame, Dawn; Gottsberger, Gerhard (21 April 2023). "Diverse sexual strategies in fossil gymnosperms: pollination in the Bennettitales revisited". Phyton. 62/63.
- S2CID 85458053.
- S2CID 134847380.
- PMID 26166781.
- ISBN 978-0-521-59283-3, retrieved 2023-03-20
- doi:10.12705/652.16.
- ^ .
- .
- ^ S2CID 135237376.
- .
- ^ Engler, H.G.A. 1892. Syllabus der Vorlesungen über specielle und medicinisch-pharmaceutische Botanik. xxiii + 184 pp. Gebr. Borntraeger, Berlin
- .
- S2CID 85709030.
- S2CID 83801635.
- PMID 29925623.
- S2CID 1198220.
- S2CID 84063572.
- S2CID 3604369.
- S2CID 234064909.
- S2CID 52253114.
- PMID 30573628.
- ^ Kustatscher, Evelyn; van Konijnenburg-van Cittert, Johanna H.A. (2005). "The Ladinian Flora (Middle Triassic) of the Dolomites: palaeoenvironmental reconstructions and palaeoclimatic considerations" (PDF). Geo.Alp. 2: 31–51.
- .
- S2CID 52885618.
- .
- ^ a b Anderson, John M.; Anderson, Heidi M.; Cleal, Chris J. (2007). "Brief history of the gymnosperms: classification, biodiversity, phytogeography and ecology" (PDF). Strelitzia. 20: 1–280.
- ^ Anderson, John M.; Anderson, Heidi M. (2003). "Heyday of the gymnosperms: systematics and biodiversity of the Late Triassic Molteno fructifications". Strelitzia. 15: 1–308.