
Choristodera
| Choristodera | |
|---|---|

| |
| Skeleton of Philydrosaurus proselius exhibited at the National Museum of Natural Science, Taiwan | |
.jpg)
| |
| Skeleton of Ikechosaurus, on display at National Museum of Natural Science, Taiwan | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | Neodiapsida |
| Order: | †Choristodera Cope, 1876 |
| Subgroups | |
| |
Choristodera (from the
History of discovery

Choristodera was erected in 1876, originally as a
Description
Choristoderes vary substantially in size, the smallest genera like
Skeletal anatomy
According to Matsumoto and colleagues (2019), choristoderes are united by the presence of 9
-
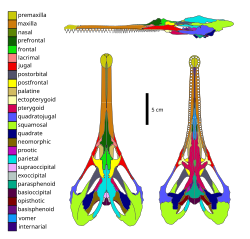 Skull diagram of Champsosaurus lindoei, a neochoristoderan
Skull diagram of Champsosaurus lindoei, a neochoristoderan -
 Skull of Lazarussuchus, an allochoristodere
Skull of Lazarussuchus, an allochoristodere


Internal skull anatomy
The internal skull anatomy of choristoderes is only known for Champsosaurus. The
Dentition
Most choristoderes have rather simple undifferentiated (
Skin


An exceptionally preserved specimen of Monjurosuchus preserves pleated skin, which indicates that in life it was probably thin and soft. The preserved scales are small and overlapping, and are smaller on the ventral underside of the body than the dorsal surface. A double row of larger ovoid scales runs along the dorsum (upper midline) of the body. The fossil also preserves webbed feet.[14] Hyphalosaurus was covered in scales of varying shape, depending on their position on the body, with at least one and possibly multiple rows of large ovoid scales running down sides of the trunk and tail. The feet display evidence of webbing, and the tail probably had additional tissue at the top and bottom, allowing it to be used as a fin to propel Hyphalosaurus through the water.[11][15] Skin impressions of Champsosaurus have also been reported, they consist of small (0.6-0.1 mm) pustulate and rhomboid scales, with the largest scales being located on the lateral sides of the body, decreasing in size dorsally, no osteoderms were present.[16] The Menat specimen of Lazarussuchus preserves some remnants of soft tissue, but no scales, which shows that the hindfoot (pes) was not webbed, and a dark stained region with a crenellated edge is present above the caudal vertebrae of the tail, suggestive of a crest similar to those found in some living reptiles, like the tuatara, lizards and crocodiles.[17]
Paleobiology
Choristoderes are exclusively found in freshwater deposits, often associated with
Diet
Neochoristoderans are presumed to have been
Reproduction
A specimen of Hyphalosaurus baitaigouensis has been found with 18 fully developed embryos within the mother's body, suggesting that they were
Tracks
Tracks from the Early Cretaceous (Albian) of South Korea, given the ichnotaxon name Novapes ulsanensis have been attributed to choristoderans, based on the similarity of the pentadactyl (five fingered) preserved tracks to the foot morphology of Monjurosuchus. The tracks preserve traces of webbing between the digits. The authors of the study proposed based on the spacing of the prints, that choristoderans could "high walk" like modern crocodilians.[25] Tracks attributed to neochoristoderans dubbed Champsosaurichnus parfeti have also been reported from the Late Cretaceous Laramie Formation of the United States, though only two prints are present and it is not possible to distinguish between a manus (forefoot) or pes (hindfoot).[26]
Classification and phylogeny

Internal systematics
Historically, the internal phylogenetics of Choristodera were unclear, with the neochoristoderes being recovered as a well-supported clade, but the relationships of the "non-neochoristoderes" being poorly resolved.[7] However, during the 2010s, the "non-neochoristoderes" from the Early Cretaceous of Asia (with the exception of Heishanosaurus) alongside Lazarussuchus from the Cenozoic of Europe were recovered (with weak support) as belonging to a monophyletic clade, which were informally named the "Allochoristoderes" by Dong and colleagues in 2020, characterised by the shared trait of completely closed lower temporal fenestrae, with Cteniogenys from the Middle-Late Jurassic of Europe and North America being consistently recovered as the basalmost choristodere.[10] The long necked "non-neochoristoderes" Shokawa and Hyphalosaurus have often been recovered as a clade, dubbed the Hyphalosauridae by Gao and Fox in 2005.[27] The finding of more complete material of the previously fragmentary Khurendukhosaurus shows that it also has a long neck, and it has also been recovered as part of the clade.[28]
Phylogeny from the analysis of Dong and colleagues (2020):[10]
| Choristodera |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Relationships to other reptiles
Choristoderes are universally agreed to be members of
Evolutionary history

Choristoderes must have diverged from all other known reptile groups prior to the end of the Permian period, over 250 million years ago, based on their primitive phylogenetic position.[5] In 2015, Rainer R. Schoch reported a new small (~ 20 cm long) diapsid from the Middle Triassic (Ladinian) Lower Keuper of Southern Germany, known from both cranial and postcranial material, which he claimed represented the oldest known choristodere.[31] Pachystropheus from the Late Triassic (Rhaetian) of Britain has been suggested to be a choristodere,[32] but cannot be referred in confidence to the group as it lacks cranial material, on which most diagnostic characters of Choristodera are based.[5] The oldest unequivocal choristoderan is the small lizard-like Cteniogenys, the oldest known remains of which are known from the late Middle Jurassic (Bathonian ~168-166 million years ago) Forest Marble and Kilmaluag formations of Britain, with remains also known from the Upper Jurassic Alcobaça Formation of Portugal and the Morrison Formation of the United States, with broadly similar remains also known from the late Middle Jurassic (Callovian) Balabansai Formation of Kyrgyzstan in Central Asia,[5][33] the Bathonian Itat Formation of western Siberia,[34] as well as possibly the Bathonian aged Anoual Formation in Morocco, North Africa.[35]
Choristoderes underwent a major evolutionary radiation in Asia during the Early Cretaceous, which represents the high point of choristoderan diversity, including the first records of the gharial-like Neochoristodera, which appear to have evolved in the regional absence of aquatic neosuchian crocodyliformes.[5] A partial femur of an indeterminate choristodere is known from the Yellow Cat Member of the Cedar Mountain Formation in North America.[36] They appear to be absent from the well sampled European localities of the Berriasian aged Purbeck Group, Great Britain and the Barremian aged La Huérguina Formation, Spain,[5] though there is a record of a small Cteniogenys-like taxon from the Berriasian aged Angeac-Charente bonebed in France.[37] In the latter half of the Late Cretaceous (Campanian-Maastrichtian), the neochoristodere Champsosaurus is found in Utah, Wyoming, Montana, North Dakota, Alberta and Saskatchewan, which were along the western coast of the Western Interior Seaway on the island of Laramidia.[5] Indeterminate remains of neochoristoderes are also known from the Canadian High Arctic, dating to the early Late Cretaceous (Coniacian–Turonian)[38] and from the Navesink Formation of New Jersey from the latest Cretaceous (Maastrichtian), which formed the separate island of Appalachia.[39] Vertebrae from the Cenomanian of Germany[40] and the Campanian aged Grünbach Formation of Austria[41] indicate the presence of choristoderes in Europe during this time period. The only record of choristoderes from Asia in the Late Cretaceous is a single vertebra from the Turonian of Japan.[42] Fragmentary remains found in the Campanian aged Oldman and Dinosaur Park formations in Alberta, Canada, also possibly suggest the presence of small bodied "non-neochoristoderes" in North America during the Late Cretaceous.
Champsosaurus survived the K-Pg extinction, and together with fellow neochoristodere Simoedosaurus are present in Europe, Asia and North America during the Paleocene, however they became extinct during the early Eocene. Their extinction coincides with major faunal turnover associated with elevated temperatures.[5] Small bodied "non-neochoristoderes", which are absent from the fossil record after the Early Cretaceous (except for possible North American remains), reappear in the form of the lizard-like Lazarussuchus from the late Paleocene of France.[17] The European endemic Lazarussuchus is the last known choristodere, surviving the extinction of neochoristoderes at the beginning of the Eocene, with the youngest known remains being those of L. dvoraki from the Early Miocene of the Czech Republic[5][43] and as well as possible indeterminate remains of Lazarussuchus reported from the late Miocene (~11.6 million years ago) of southern Germany.[44]
References
- ^ PMID 35313822.
- ISBN 978-1-4214-2867-3.
- ^ Cope ED (1876). "On some extinct reptiles and Batrachia from the Judith River and Fox Hills beds of Montana". Proceedings of the Academy of Natural Sciences of Philadelphia. 1876: 340–359.
- ^ Cope, ED (1884). "The Choristodera". The American Naturalist. 18: 815–817.
- ^ .
- ^ ISBN 978-1-4615-2878-4.
- ^ S2CID 92421503.
- ^ PMID 31905243.
- ISSN 2193-0074.
- ^ S2CID 219047160.
- ^ PMID 18510504.
- PMID 32346021.
- PMID 26573112.
- ^ S2CID 86064702.
- .
- ^ .
- ^ S2CID 129438118.
- S2CID 244917425.
- S2CID 95194181.
- ^ PMC 10333127.
- S2CID 8719805.
- PMID 20018793.
- ^ S2CID 49570935.
- S2CID 84966652.
- PMID 32879388.
- ISSN 0272-4634.
- ISSN 1096-3642.
- S2CID 133925482.
- ^ PMID 27162705.
- .
- ^ Schoch, R.R (2015). "Reptilien" (PDF). Der Lettenkeuper: ein Fenster in die Zeit vor den Dinosauriern (in German). Staatliches Museum für Naturkunde Stuttgart. pp. 231–264.
- S2CID 86088809.
- S2CID 7111980.
- .
- .
- S2CID 86258448.
- ^ Ronan Allain, Romain Vullo, Lee Rozada, Jérémy Anquetin, Renaud Bourgeais, et al.. Vertebrate paleobiodiversity of the Early Cretaceous (Berriasian) Angeac-Charente Lagerstätte (southwestern France): implications for continental faunal turnover at the J/K boundary. Geodiversitas, Museum National d’Histoire Naturelle Paris, In press. ffhal-03264773f
- .
- S2CID 237761128.
- S2CID 134904339.
- ^ Buffetaut, Eric (1989). "Erster nachweis von Choristodera (Reptilia, Diapsida) in der Oberkreide Europas: Champsosaurierwirbel aus den Gosau-Schichten (Campan) Niederösterreichs" [First evidence of Choristodera (Reptilia, Diapsida) in the Upper Cretaceous of Europe: Champsosaur vertebrae from the Gosau strata (Campanian) of Lower Austria] (PDF). Sitzungsberichten der Österreichs Akademis der Wissenschaften Mathematisch-Naturwissenschaftlichen Klasse, Abteilung (in German). 197: 389–394.
- S2CID 238680387.
- S2CID 84097919.
- ISSN 0078-0421.
Further reading
- de Braga M, Rieppel O (1997). "Reptile phylogeny and the interrelationships of turtles". Zoological Journal of the Linnean Society. 120 (3): 281–354. .
- Erickson BR (1972). The Lepidosaurian Reptile Champsosaurus in North America. Paleontology, Monograph. Vol. 1. Science Museum of Minnesota.
- Matsumoto R, Suzuki S, Tsogtbaatar K, Evans SE (February 2009). "New material of the enigmatic reptile Khurendukhosaurus (Diapsida: Choristodera) from Mongolia". Die Naturwissenschaften. 96 (2): 233–42. S2CID 13542692.
- Ksepka D, Gao K, Norell MA (2005). "A new choristodere from the Cretaceous of Mongolia". American Museum Novitates (3468): 1–22. S2CID 54179348.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||







| Authority control databases: National |
|---|