
Echinoderm
| Echinoderms | |
|---|---|

| |
| Extant and extinct echinoderms of six classes: Echinoidea); Oxycomanthus bennetti (Crinoidea); Actinopyga echinites (Holothuroidea); Ctenocystoidea .
| |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Subkingdom: | Eumetazoa |
| Clade: | ParaHoxozoa |
| Clade: | Bilateria |
| Clade: | Nephrozoa |
| Superphylum: | Deuterostomia |
| Clade: | Ambulacraria |
| Phylum: | Echinodermata Bruguière, 1791 [ex Klein, 1734] |
| Type genus | |
| Echinus Linnaeus, 1758
| |
| Subphyla and classes[2] | |
|
Homalozoa † Gill & Caster, 1960
†=Extinct | |
An echinoderm (
The echinoderms are important both ecologically and geologically. Ecologically, there are few other groupings so abundant in the
Taxonomy and evolution
The name echinoderm is from
Phylogeny
Historically, taxonomists believed that the Ophiuroidea were sister to the Asteroidea, or that they were sister to the (Holothuroidea + Echinoidea).[9] However, a 2014 analysis of 219 genes from all classes of echinoderms revised the phylogenetic tree.[10] An independent analysis in 2015 of RNA transcriptomes from 23 species across all classes of echinoderms gave the same tree.[9]
- External phylogeny
The context of the echinoderms within the Bilateria is:[11]
| Bilateria |
| ||||||||||||||||||||||||
_001.png)
- Internal phylogeny
| Echino‑ | |
| dermata |


.jpg)
Diversity
There are about 7,000
All echinoderms are- The subphyla of echinoderms
-
 A brittle star, Ophionereis reticulata
A brittle star, Ophionereis reticulata -
 A sea cucumber, Stichopus chloronotus, from Malaysia
A sea cucumber, Stichopus chloronotus, from Malaysia -
 Starfish of varied colours
Starfish of varied colours -
 A sea urchin, Strongylocentrotus purpuratus
A sea urchin, Strongylocentrotus purpuratus -
 Crinoid on a coral reef
Crinoid on a coral reef





Fossil history
The oldest candidate echinoderm fossil is Arkarua from the Precambrian of Australia. These fossils are disc-like, with radial ridges on the rim and a five-pointed central depression marked with radial lines. However, the fossils have no stereom or internal structure indicating a water vascular system, so they cannot be conclusively identified.[15]
The first universally accepted echinoderms appear in the
The first echinoderms were non-motile,
- Further information: Dibrachicystis
-
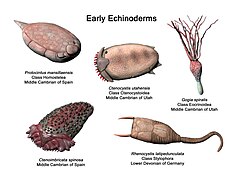
-
 The Ordovician cystoid Echinosphaerites from northeastern Estonia
The Ordovician cystoid Echinosphaerites from northeastern Estonia -
 Fossil crinoid crowns
Fossil crinoid crowns -





Anatomy and physiology
Echinoderms evolved from animals with
Echinoderms have secondary radial symmetry in portions of their body at some stage of life, most likely an adaptation to a sessile or slow-moving existence.
Genetic studies have showed that genes which direct the development of the trunk region is absent during the development (only known exception is a gene active at the tip of the sea stars arms), indicating that their body consists of just a developed head region.[23][24]
Skin and skeleton
Echinoderms have a
Although individual ossicles are robust and fossilize readily, complete skeletons of starfish, brittle stars and crinoids are rare in the fossil record. On the other hand, sea urchins are often well preserved in chalk beds or limestone. During fossilization, the cavities in the stereom are filled in with calcite that is continuous with the surrounding rock. On fracturing such rock, paleontologists can observe distinctive cleavage patterns and sometimes even the intricate internal and external structure of the test.[27]
The epidermis contains pigment cells that provide the often vivid colours of echinoderms, which include deep red, stripes of black and white, and intense purple.[28] These cells may be light-sensitive, causing many echinoderms to change appearance completely as night falls. The reaction can happen quickly: the sea urchin Centrostephanus longispinus changes colour in just fifty minutes when exposed to light.[29]
One characteristic of most echinoderms is a special kind of tissue known as catch connective tissue. This collagen-based material can change its mechanical properties under nervous control rather than by muscular means. This tissue enables a starfish to go from moving flexibly around the seabed to becoming rigid while prying open a bivalve mollusc or preventing itself from being extracted from a crevice. Similarly, sea urchins can lock their normally mobile spines upright as a defensive mechanism when attacked.[30][31]
The water vascular system

Echinoderms possess a unique water vascular system, a network of fluid-filled canals modified from the
The organisation of the water vascular system is somewhat different in ophiuroids, where the madreporite may be on the oral surface and the podia lack suckers.[34] In holothuroids, the system is reduced, often with few tube feet other than the specialised feeding tentacles, and the madreporite opens on to the coelom. Some holothuroids like the Apodida lack tube feet and canals along the body; others have longitudinal canals.[35] The arrangement in crinoids is similar to that in asteroids, but the tube feet lack suckers and are used in a back-and-forth wafting motion to pass food particles captured by the arms towards the central mouth. In the asteroids, the same motion is employed to move the animal across the ground.[36]
Other organs
Echinoderms possess a simple digestive system which varies according to the animal's diet. Starfish are mostly carnivorous and have a mouth, oesophagus, two-part stomach, intestine and rectum, with the anus located in the centre of the aboral body surface. With a few exceptions, the members of the order
The
Haemal and perihaemal systems are derived from the original coelom, forming an
The coelomic fluid contains the
Echinoderms have a simple radial
The gonads at least periodically occupy much of the body cavities of sea urchins[52] and sea cucumbers, while the less voluminous crinoids, brittle stars and starfish have two gonads in each arm. While the ancestors of modern echinoderms are believed to have had one genital aperture, many organisms have multiple gonopores through which eggs or sperm may be released.[51]
Regeneration
Many echinoderms have great powers of
The regeneration of lost parts involves both epimorphosis and morphallaxis. In epimorphosis stem cells—either from a reserve pool or those produced by dedifferentiation—form a blastema and generate new tissues. Morphallactic regeneration involves the movement and remodelling of existing tissues to replace lost parts.[58] Direct transdifferentiation of one type of tissue to another during tissue replacement is also observed.[59]
Reproduction
Sexual reproduction
Echinoderms become sexually mature after approximately two to three years, depending on the species and the environmental conditions. Almost all species have separate
Some echinoderms brood their eggs. This is especially common in cold water species where planktonic larvae might not be able to find sufficient food. These retained eggs are usually few in number and are supplied with large yolks to nourish the developing embryos. In starfish, the female may carry the eggs in special pouches, under her arms, under her arched body, or even in her cardiac stomach.[61] Many brittle stars are hermaphrodites; they often brood their eggs, usually in special chambers on their oral surfaces, but sometimes in the ovary or coelom.[62] In these starfish and brittle stars, development is usually direct to the adult form, without passing through a bilateral larval stage.[63] A few sea urchins and one species of sand dollar carry their eggs in cavities, or near their anus, holding them in place with their spines.[64] Some sea cucumbers use their buccal tentacles to transfer their eggs to their underside or back, where they are retained. In a very small number of species, the eggs are retained in the coelom where they develop viviparously, later emerging through ruptures in the body wall.[65] In some crinoids, the embryos develop in special breeding bags, where the eggs are held until sperm released by a male happens to find them.[66]
Asexual reproduction

One species of
Adult sea cucumbers reproduce asexually by transverse fission. Holothuria parvula uses this method frequently, splitting into two a little in front of the midpoint. The two halves each regenerate their missing organs over a period of several months, but the missing genital organs are often very slow to develop.[70]
The larvae of some echinoderms are capable of asexual reproduction. This has long been known to occur among starfish and brittle stars, but has more recently been observed in a sea cucumber, a sand dollar and a sea urchin.
Cloning is costly to the larva both in resources and in development time. Larvae undergo this process when food is plentiful[74] or temperature conditions are optimal.[73] Cloning may occur to make use of the tissues that are normally lost during metamorphosis.[75] The larvae of some sand dollars clone themselves when they detect dissolved fish mucus, indicating the presence of predators.[73][75] Asexual reproduction produces many smaller larvae that escape better from planktivorous fish, implying that the mechanism may be an anti-predator adaptation.[76]
Larval development
Development begins with a bilaterally symmetrical embryo, with a coeloblastula developing first. Gastrulation marks the opening of the "second mouth" that places echinoderms within the deuterostomes, and the mesoderm, which will host the skeleton, migrates inwards. The secondary body cavity, the coelom, forms by the partitioning of three body cavities. The larvae are often planktonic, but in some species the eggs are retained inside the female, while in some the female broods the larvae.[77][78]
The larvae pass through several stages, which have specific names derived from the taxonomic names of the adults or from their appearance. For example, a sea urchin has an 'echinopluteus' larva while a brittle star has an 'ophiopluteus' larva. A starfish has a '
A
Distribution and habitat
Echinoderms are globally distributed in almost all depths, latitudes and environments in the ocean. Adults are mainly benthic, living on the seabed, whereas larvae are often pelagic, living as plankton in the open ocean. Some holothuroid adults such as Pelagothuria are however pelagic.[80] Some crinoids are pseudo-planktonic, attaching themselves to floating logs and debris, although this behaviour was exercised most extensively in the Paleozoic, before competition from organisms such as barnacles restricted the extent of the behaviour.[81]
Mode of life
Locomotion

Echinoderms primarily use their tube feet to move about, though some sea urchins also use their spines. The tube feet typically have a tip shaped like a suction pad in which a vacuum can be created by contraction of muscles. This combines with some stickiness from the secretion of mucus to provide adhesion. The tube feet contract and relax in waves which move along the adherent surface, and the animal moves slowly along.[82]
Brittle stars are the most agile of the echinoderms. Any one of the arms can form the axis of symmetry, pointing either forwards or back. The animal then moves in a co-ordinated way, propelled by the other four arms. During locomotion, the propelling arms can made either snake-like or rowing movements.
Sea urchins use their tube feet to move around in a similar way to starfish. Some also use their articulated spines to push or lever themselves along or lift their oral surfaces off the substrate. If a sea urchin is overturned, it can extend its tube feet in one ambulacral area far enough to bring them within reach of the substrate and then successively attach feet from the adjoining area until it is righted. Some species bore into rock, usually by grinding away at the surface with their mouthparts.[86]
.jpg)
Sea cucumbers are generally sluggish animals. Many can move on the surface of the seabed or burrow through sand or mud using
The majority of crinoids are motile, but sea lilies are sessile and attached to hard substrates by stalks. Movement in most sea lilies is limited to bending their stems can bend and rolling and unrolling their arms; a few species can relocate themselves on the seabed by crawling. The sea feathers are unattached and usually live in crevices, under corals or inside sponges with their arms the only visible part. Some sea feathers emerge at night and perch themselves on nearby eminences to better exploit food-bearing currents. Many species can "walk" across the seabed, raising their body with the help of their arms, or swim using their arms. Most species of sea feather, however, are largely sedentary, seldom moving far from their chosen place of concealment.[88]
Feeding
The modes of feeding vary greatly between the different echinoderm taxa. Crinoids and some brittle stars tend to be passive filter-feeders,[89][90] enmeshing suspended particles from passing water. Most sea urchins are grazers;[91] sea cucumbers are deposit feeders;[92] and the majority of starfish are active hunters.[93]
Crinoids catch food particles using the tube feet on their outspread pinnules, move them into the ambulacral grooves, wrap them in mucus, and convey them to the mouth using the cilia lining the grooves.[89] The exact dietary requirements of crinoids have been little researched, but in the laboratory, they can be fed with diatoms.[94]
Basket stars are suspension feeders, raising their branched arms to collect zooplankton, while other brittle stars use several methods of feeding. Some are suspension feeders, securing food particles with mucus strands, spines or tube feet on their raised arms. Others are scavengers and detritus feeders. Others again are voracious carnivores and able to lasso their waterborne prey with a sudden encirclement by their flexible arms. The limbs then bend under the disc to transfer the food to the jaws and mouth.[90]
Many sea urchins feed on algae, often scraping off the thin layer of algae covering the surfaces of rocks with their specialised mouthparts known as Aristotle's lantern. Other species devour smaller organisms, which they may catch with their tube feet. They may also feed on dead fish and other animal matter.[91] Sand dollars may perform suspension feeding and feed on phytoplankton, detritus, algal pieces and the bacterial layer surrounding grains of sand.[95]
Sea cucumbers are often mobile deposit or suspension feeders, using their buccal podia to actively capture food and then stuffing the particles individually into their buccal cavities. Others ingest large quantities of sediment, absorb the organic matter and pass the indigestible mineral particles through their guts. In this way they disturb and process large volumes of substrate, often leaving characteristic ridges of sediment on the seabed. Some sea cucumbers live infaunally in burrows, anterior-end down and anus on the surface, swallowing sediment and passing it through their gut. Other burrowers live anterior-end up and wait for detritus to fall into the entrances of the burrows or rake in debris from the surface nearby with their buccal podia.[92]
Nearly all starfish are detritus feeders or carnivores, though a few are suspension feeders. Small fish landing on the upper surface may be captured by pedicilaria and dead animal matter may be scavenged but the main prey items are living invertebrates, mostly bivalve molluscs. To feed on one of these, the starfish moves over it, attaches its tube feet and exerts pressure on the valves by arching its back. When a small gap between the valves is formed, the starfish inserts part of its stomach into the prey, excretes digestive enzymes and slowly liquefies the soft body parts. As the adductor muscle of the bivalve relaxes, more stomach is inserted and when digestion is complete, the stomach is returned to its usual position in the starfish with its now liquefied bivalve meal inside it. Other starfish evert the stomach to feed on sponges, sea anemones, corals, detritus and algal films.[93]
Antipredator defence
.jpg)
Despite their low nutrition value and the abundance of indigestible calcite, echinoderms are preyed upon by many organisms, including
Ecology

Echinoderms are numerous invertebrates whose adults play an important role in benthic ecosystems, while the larvae are a major component of the plankton. Among the ecological roles of adults are the grazing of sea urchins, the sediment processing of heart urchins, and the suspension and deposit feeding of crinoids and sea cucumbers.[13][80] Some sea urchins can bore into solid rock, destabilising rock faces and releasing nutrients into the ocean. Coral reefs are also bored into in this way, but the rate of accretion of carbonate material is often greater than the erosion produced by the sea urchin.[100] Echinoderms sequester about 0.1 gigatonnes of carbon dioxide per year as calcium carbonate, making them important contributors in the global carbon cycle.[101]
Echinoderms sometimes have large population swings which can transform ecosystems. In 1983, for example, the mass mortality of the tropical sea urchin Diadema antillarum in the Caribbean caused a change from a coral-dominated reef system to an alga-dominated one.[102] Sea urchins are among the main herbivores on reefs and there is usually a fine balance between the urchins and the kelp and other algae on which they graze. A diminution of the numbers of predators (otters, lobsters and fish) can result in an increase in urchin numbers, causing overgrazing of kelp forests, resulting in an alga-denuded "urchin barren".[103] On the Great Barrier Reef, an unexplained increase in the numbers of crown-of-thorns starfish (Acanthaster planci), which graze on living coral tissue, has greatly increased coral mortality and reduced coral reef biodiversity.[104]
Use by humans
As food and medicine


In 2019, 129,052 tonnes of echinoderms were harvested. The majority of these were sea cucumbers (59,262 tonnes) and sea urchins (66,341 tonnes).[105] These are used mainly for food, but also in traditional Chinese medicine.[106] Sea cucumbers are considered a delicacy in some countries of southeast Asia; as such, they are in imminent danger of being over-harvested.[107] Popular species include the pineapple roller Thelenota ananas (susuhan) and the red sea cucumber Holothuria edulis. These and other species are colloquially known as bêche de mer or trepang in China and Indonesia. The sea cucumbers are boiled for twenty minutes and then dried both naturally and later over a fire which gives them a smoky tang. In China they are used as a basis for gelatinous soups and stews.[108] Both male and female gonads of sea urchins are consumed, particularly in Japan and France. The taste is described as soft and melting, like a mixture of seafood and fruit.[109][110] Sea urchin breeding trials have been undertaken to try to compensate for overexploitation.[111]
In research
Because of their robust larval growth, sea urchins are widely used in research, particularly as
Other uses
The calcareous tests or shells of echinoderms are used as a source of
See also
References
- ^ Smith, A., Zamora, S. & Álvaro, J. The oldest echinoderm faunas from Gondwana show that echinoderm body plan diversification was rapid. Nat Commun 4, 1385 (2013). https://doi.org/10.1038/ncomms2391.
- ^ Stöhr, Sabine (2014). "Echinodermata". WoRMS. World Register of Marine Species. Retrieved 23 February 2014.
- ^ "echinoderm". Dictionary.com Unabridged (Online). n.d.
- ^ Hall, Danielle (3 February 2022). "Echinoderms | Smithsonian Ocean". ocean.si.edu. Retrieved 31 March 2023.
- ^ "echinoderm". Online Etymology Dictionary.
- ^ Dorit, Walker & Barnes 1991, pp. 777–779.
- ^ Fox, Richard. "Asterias forbesi". Invertebrate Anatomy OnLine. Lander University. Retrieved 19 May 2012.
- ^ a b Wray, Gregory A. (1999). "Echinodermata: Spiny-skinned animals: sea urchins, starfish, and their allies". Tree of Life. Archived from the original on 23 May 2015. Retrieved 19 October 2012.
- ^ PMID 25794146.
- PMID 24850925.
- S2CID 32169826.
- ^ "Animal Diversity Web - Echinodermata". University of Michigan Museum of Zoology. Retrieved 26 August 2012.
- ^ .
- ISBN 978-3-7091-1855-9.
- ^ Smith, Dave (28 September 2005). "Vendian Animals: Arkarua". Retrieved 14 March 2013.
- ^ Waggoner, Ben (16 January 1995). "Echinodermata: Fossil Record". Introduction to the Echinodermata. Museum of Paleontology: University of California at Berkeley. Retrieved 14 March 2013.
- ^ a b c Dorit, Walker & Barnes 1991, pp. 792–793.
- ^ UCMP Berkeley. "Echinodermata: Morphology". University of California Museum of Paleontology. Retrieved 21 March 2011.
- ^ a b Ruppert, Fox & Barnes 2004, p. 873.
- )
- PMID 22789130.
- ^ Messing, Charles. "Crown and calyx". Charles Messing's Crinoid Pages. Archived from the original on 29 October 2013. Retrieved 29 July 2012.
- ^ Molecular evidence of anteroposterior patterning in adult echinoderms
- ^ New Genetic Study Reveals Sea Stars Are Just A Bunch of Heads Smushed Together
- ISBN 978-3-527-31805-6.
- ^ Brusca, Moore & Shuster 2016, pp. 979–980.
- ^ Davies, A. Morley (1925). An Introduction to Palaeontology. Thomas Murby. pp. 240–241.
- PMID 32812865.
- S2CID 1669387.
- S2CID 86391676.
- ^ Brusca, Moore & Shuster 2016, p. 980.
- ^ a b Dorit, Walker & Barnes 1991, pp. 780–791.
- ^ Brusca, Moore & Shuster 2016, pp. 980–982.
- ^ Dorit, Walker & Barnes 1991, pp. 784–785.
- ^ Brusca, Moore & Shuster 2016, p. 982.
- ^ Dorit, Walker & Barnes 1991, pp. 790–793.
- ISBN 978-90-6191-080-0.
- ISBN 978-9971693688.
- ^ Ruppert, Fox & Barnes 2004, p. 885.
- ^ Ruppert, Fox & Barnes 2004, p. 891.
- ^ Ruppert, Fox & Barnes 2004, pp. 902–904.
- ^ Ruppert, Fox & Barnes 2004, p. 912.
- ^ Ruppert, Fox & Barnes 2004, p. 920.
- ISBN 978-0521674065.
- OCLC 1148032077.
- ^ "Macrobenthos of the North Sea - Echinodermata > Introduction". etibioinformatics.nl.
- ISBN 978-0199606023.
- ^ a b Ramirez-Gomez, Fransisco (27 September 2010). "Echinoderm Immunity". Invertebrate Survival Journal.
- PMID 21528703.
- PMID 21528703.
- ^ a b Ruppert, Fox & Barnes 2004, pp. 872–929.
- ^ James, Philip; Siikavuopio, Sten; Johansson, Gunhild Seljehaug (2018). "A Guide to the Sea Urchin Reproductive Cycle and Staging Sea Urchin Gonad Samples" (PDF). Nofima. Archived (PDF) from the original on 9 October 2022. Retrieved 21 June 2022.
The GI [% of body weight made up by the gonad] of urchins in the wild can vary hugely and can be less than 1% or as high as 20%, whilst for cultured sea urchins GI values can be as high as 35%
- ^ Edmondson, C.H. (1935). "Autotomy and regeneration of Hawaiian starfishes" (PDF). Bishop Museum Occasional Papers. 11 (8): 3–20. Archived (PDF) from the original on 9 October 2022.
- ^ a b McAlary, Florence A. (1993). Population Structure and Reproduction of the Fissiparous Seastar, Linckia columbiae Gray, on Santa Catalina Island, California. 3rd California Islands Symposium. National Park Service. Retrieved 15 April 2012.
- ^ .
- ^ (PDF) from the original on 9 October 2022. Retrieved 15 July 2011.
- PMID 29303627. Retrieved 14 July 2011.
- ISBN 978-90-481-2766-5.
- S2CID 27847466.
- ISBN 0231080042.
- ^ Ruppert, Fox & Barnes 2004, pp. 887–888.
- ^ Ruppert, Fox & Barnes 2004, p. 895.
- ^ Ruppert, Fox & Barnes 2004, p. 888.
- ^ Ruppert, Fox & Barnes 2004, p. 908.
- ^ Ruppert, Fox & Barnes 2004, p. 916.
- ^ Ruppert, Fox & Barnes 2004, p. 922.
- S2CID 84475593.
- (PDF) from the original on 9 October 2022.
- JSTOR 4063000.
- (PDF) from the original on 9 October 2022.
- ^ S2CID 4430104.
- (PDF) from the original on 9 October 2022. Retrieved 13 July 2011.
- ^ S2CID 26030855.
- S2CID 24422314. Retrieved 16 July 2011.
- ^ S2CID 206509992. Retrieved 16 July 2011.
- S2CID 84336247.
- ^ a b Dorit, Walker & Barnes 1991, p. 778.
- ^ Brusca, Moore & Shuster 2016, pp. 997–998.
- ^ van Egmond, Wim (1 July 2000). "Gallery of Echinoderm Larvae". Microscopy UK. Retrieved 2 February 2013.
- ^ a b Brusca, Moore & Shuster 2016, p. 968.
- .
- S2CID 55933156. Archived from the original(PDF) on 15 November 2013.
- PMID 22573771.
- ^ Brusca, Moore & Shuster 2016, pp. 982–983.
- ^ "Sand star - Luidia foliolata". Sea Stars of the Pacific Northwest. Archived from the original on 9 September 2012. Retrieved 26 September 2012.
- ^ Ruppert, Fox & Barnes 2004, pp. 899–900.
- ^ Ruppert, Fox & Barnes 2004, pp. 911–912.
- ^ Messing, Charles. "The crinoid feeding mechanism". Charles Messing's Crinoid Pages. Archived from the original on 14 January 2013. Retrieved 26 July 2012.
- ^ ISBN 0-03-056747-5.
- ^ a b Ruppert, Fox & Barnes 2004, p. 893.
- ^ a b Carefoot, Tom. "Learn about sea urchins: feeding, nutrition & growth". A Snail's Odyssey. Archived from the original on 22 March 2013. Retrieved 23 February 2013.
- ^ a b Ruppert, Fox & Barnes 2004, p. 914.
- ^ a b Ruppert, Fox & Barnes 2004, pp. 884–885.
- ^ Carefoot, Tom. "Learn about feather stars: feeding & growth". A Snail's Odyssey. Archived from the original on 12 January 2013. Retrieved 23 February 2013.
- ^ Carefoot, Tom. "Learn about sand dollars: feeding & growth". A Snail's Odyssey. Archived from the original on 19 October 2012. Retrieved 23 February 2013.
- ^ Miller, John E. "Echinoderm: role in nature". Encyclopædia Britannica. Retrieved 23 June 2022.
- ^ Dorit, Walker & Barnes 1991, p. 779.
- ^ Dorit, Walker & Barnes 1991, pp. 789–790.
- JSTOR 1541585.
- PMID 17469255.
- hdl:10261/79231.
- ISBN 0-521-64523-9.
- ^ Lawrence, J. M. (1975). "On the relationships between marine plants and sea urchins" (PDF). Oceanographic Marine Biological Annual Review. 13: 213–286. Archived from the original (PDF) on 5 February 2011.
- ISBN 0849365996.
- ^ "B-76 Sea-urchins and other echinoderms Capture production by species, fishing areas and countries or areas" (PDF). FAO. pp. 463–465. Archived (PDF) from the original on 9 October 2022. Retrieved 23 June 2022.
- PMID 29992105.
- ^ "Sea Cucumbers Threatened by Asian Trade". The New York Times. April 2009. Retrieved 1 April 2009.
- ^ Wikisource:1911 Encyclopædia Britannica/Bêche-de-Mer
- ISBN 978-0444503909.
- Oxford Companion to Food. Oxford University Press, 3rd edition. p. 730.
- S2CID 18856183.
- ^ a b "Insight from the Sea Urchin". Microscope Imaging Station. Exploratorium. Archived from the original on 5 February 2018. Retrieved 23 June 2022.
- S2CID 207437380.
- S2CID 207437380.
- S2CID 39175905.
- PMID 5816822.
- ^ a b Barkhouse, C.; Niles, M.; Davidson, L. A. (2007). "A literature review of sea star control methods for bottom and off bottom shellfish cultures" (PDF). Canadian Industry Report of Fisheries and Aquatic Sciences. 279 (7): 14–15. Archived from the original (PDF) on 22 May 2013.
- PMID 34791383.
- ^ "Sea stars". Wild Singapore. Retrieved 4 February 2013.
Cited texts
- Brusca, Richard C.; Moore, Wendy; Shuster, Stephen M. (2016). Invertebrates (3rd ed.). Sunderland, Massachusetts. )
- Dorit, R. L.; Walker, W. F.; Barnes, R. D. (1991). Zoology, International edition. Saunders College Publishing. ISBN 978-0-03-030504-7.
- Ruppert, Edward E.; Fox, Richard, S.; Barnes, Robert D. (2004). Invertebrate Zoology (7th ed.). Cengage Learning. ISBN 81-315-0104-3.)
{{cite book}}: CS1 maint: multiple names: authors list (link
External links
- The Echinoid Directory from the Natural History Museum
- Echinodermata from the Tree of Life Web Project
- Echinoderms of the North Sea Archived 13 April 2008 at the Wayback Machine
- Larval Echinodermata Fact Sheet