
Fish anatomy

Fish anatomy is the study of the form or morphology of fish. It can be contrasted with fish physiology, which is the study of how the component parts of fish function together in the living fish.[1] In practice, fish anatomy and fish physiology complement each other, the former dealing with the structure of a fish, its organs or component parts and how they are put together, such as might be observed on the dissecting table or under the microscope, and the latter dealing with how those components function together in living fish.
The anatomy of fish is often shaped by the physical characteristics of water, the medium in which fish live. Water is much
The bony fish lineage shows more
Body

In many respects, fish anatomy is different from
Fish have a variety of different body plans. At the broadest level, their body is divided into head, trunk, and tail, although the divisions are not always externally visible. The body is often fusiform, a streamlined body plan often found in fast-moving fish. They may also be filiform (eel-shaped) or vermiform (worm-shaped). Fish are often either compressed (laterally thin) or depressed (dorso-ventrally flat).
Skeleton
There are two different skeletal types: the exoskeleton, which is the stable outer shell of an organism, and the endoskeleton, which forms the support structure inside the body. The skeleton of the fish is made of either cartilage (cartilaginous fishes) or bone (bony fishes). The endoskeleton of the fish is made up of two main components: the axial skeleton consisting of the skull and vertebral column, and the appendicular skeleton supporting the fins.[7] The fins are made up of bony fin rays and, except for the caudal fin, have no direct connection with the spine. They are supported only by the muscles. The ribs attach to the spine.
Bones are rigid
Vertebrae


Fish are vertebrates. All vertebrates are built along the basic
The defining characteristic of a vertebrate is the vertebral column, in which the notochord (a stiff rod of uniform composition) found in all chordates has been replaced by a segmented series of stiffer elements (vertebrae) separated by mobile joints (
The vertebral column consists of a
The vertebrae of
In most
In cartilaginous fish such as sharks, the vertebrae consist of two cartilaginous tubes. The upper tube is formed from the vertebral arches, but also includes additional cartilaginous structures filling in the gaps between the vertebrae, enclosing the spinal cord in an essentially continuous sheath. The lower tube surrounds the notochord and has a complex structure, often including multiple layers of calcification.[11]
Head
_skull_labeled.png)

.png)
The head or
The skull in fishes is formed from a series of only loosely connected bones. Jawless fish and sharks only possess a cartilaginous
In lampreys, the mouth is formed into an oral disk. In most jawed fish, however, there are three general configurations. The mouth may be on the forward end of the head (terminal), may be upturned (superior), or may be turned downwards or on the bottom of the fish (subterminal or inferior). The mouth may be modified into a suckermouth adapted for clinging onto objects in fast-moving water.
The simpler structure is found in jawless fish, in which the cranium is represented by a trough-like basket of cartilaginous elements only partially enclosing the brain and associated with the capsules for the inner ears and the single nostril. Distinctively, these fish have no jaws.[15]
Cartilaginous fish such as sharks also have simple, and presumably primitive, skull structures. The cranium is a single structure forming a case around the brain, enclosing the lower surface and the sides, but always at least partially open at the top as a large
In the ray-finned fishes, there has also been considerable modification from the primitive pattern. The roof of the skull is generally well formed, and although the exact relationship of its bones to those of tetrapods is unclear, they are usually given similar names for convenience. Other elements of the skull, however, may be reduced; there is little cheek region behind the enlarged orbits, and little if any bone in between them. The upper jaw is often formed largely from the
Although the skulls of fossil lobe-finned fish resemble those of the early tetrapods, the same cannot be said of those of the living lungfishes. The skull roof is not fully formed, and consists of multiple, somewhat irregularly shaped bones with no direct relationship to those of tetrapods. The upper jaw is formed from the pterygoid bones and vomers alone, all of which bear teeth. Much of the skull is formed from cartilage, and its overall structure is reduced.[15]
The head may have several fleshy structures known as
-
 Skull of a northern pike
Skull of a northern pike -
 Skull ofDevonian period
Skull ofDevonian period


External organs
Jaw
The vertebrate jaw probably originally evolved in the

| External videos | |
|---|---|
It is thought that the original selective advantage garnered by the jaw was not related to feeding, but to increase
Eyes
Fish eyes are similar to
Gills

The gills, located under the operculum, are a respiratory organ for the extraction of oxygen from water and for the excretion of carbon dioxide. They are not usually visible, but can be seen in some species, such as the
Skin
The skin of the fish are a part of the integumentary system, which contains two layers: the epidermis and the dermis layer. The epidermis is derived from the ectoderm and becomes the most superficial layer that consists entirely of live cells, with only minimal quantities of keratin. It is generally permeable. The dermis is derived from the mesoderm and resembles the little connective tissue which are composed of mostly collagen fibers found in bony fish. Some fish species have scales that emerge from the dermis, penetrate the thin layer of the basement membrane that lies between the epidermis and dermis, and becomes externally visible and covers the epidermis layer.[20]
Generally, the skin also contains

Scales
Also part of the fish's integumentary system are the scales that cover the outer body of many jawed fish. The commonly known scales are the ones that originate from the dermis or mesoderm, and may be similar in structure to teeth. Some species are covered by scutes instead. Others may have no scales covering the outer body.
-
 Singular bowfin cycloid scale
Singular bowfin cycloid scale -
 Cycloid scales covering rohu
Cycloid scales covering rohu -
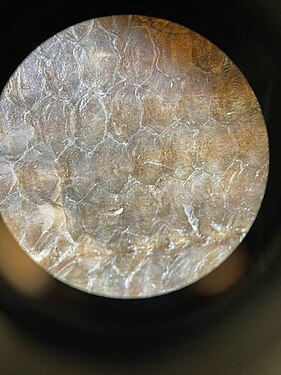 Bowfin cycloid scales
Bowfin cycloid scales



There are four principal types of fish scales that originate from the dermis.[25][26]
- have it on claspers only).
- .
- have cycloid scales.
- Ctenoid scales are similar to cycloid scales, also having growth rings, lack enamel, dentin, and a vascular bone layer. They are distinguished by spines or projections along one edge. Halibuthave this type of scale.
-
 Fish scales: 1. cycloid scale; 2. ctenoid scale; 3. placcoid scale; 4. ganoid scale
Fish scales: 1. cycloid scale; 2. ctenoid scale; 3. placcoid scale; 4. ganoid scale -
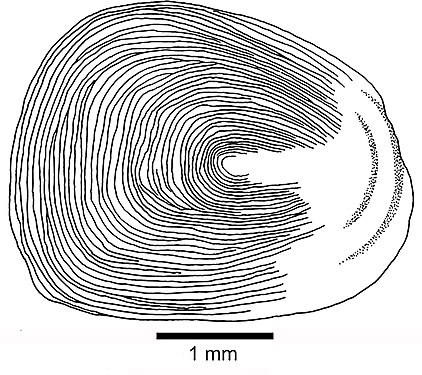 Cycloid scale
Cycloid scale -
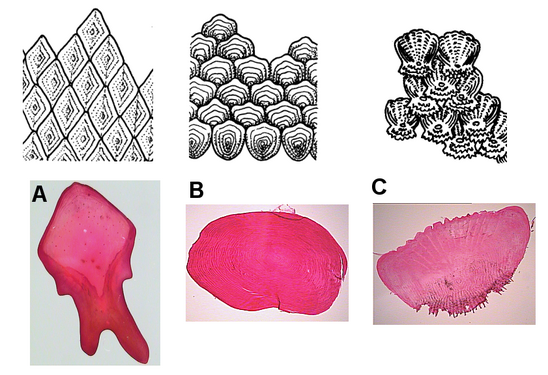 Fish scales: A. ganoid; B. cycloid; C. ctenoid
Fish scales: A. ganoid; B. cycloid; C. ctenoid
_(20544399878).jpg)


Another less common type of scale is the
Lateral line

The lateral line is a
Photophores
Photophores are light-emitting organs which appear as luminous spots on some fishes. The light can be produced from compounds during the digestion of prey, from specialized
Fins

_diagram_cropped.GIF)
Fins are the most distinctive features of fish. They are either composed of bony spines or rays protruding from the body with skin covering them and joining them together, either in a webbed fashion as seen in most bony fish, or similar to a flipper as seen in sharks. Apart from the tail or caudal fin, fins have no direct connection with the spine and are supported by muscles only. Their principal function is to help the fish swim. Fins can also be used for gliding or crawling, as seen in the flying fish and frogfish. Fins located in different places on the fish serve different purposes, such as moving forward, turning, and keeping an upright position. For every fin, there are a number of fish species in which this particular fin has been lost during evolution.[citation needed]
Spines and rays
In bony fish, most fins may have spines or rays. A fin may contain only spiny rays, only soft rays, or a combination of both. If both are present, the spiny rays are always anterior. Spines are generally stiff, sharp and unsegmented. Rays are generally soft, flexible, segmented, and may be branched. This segmentation of rays is the main difference that distinguishes them from spines; spines may be flexible in certain species, but never segmented.
Spines have a variety of uses. In catfish, they are used as a form of defense; many catfish have the ability to lock their spines outwards. Triggerfish also use spines to lock themselves in crevices to prevent them being pulled out.
Types of fin
- pterygiophores. In spinous fins, the distal pterygiophore is often fused to the middle or not present at all.
- Caudal/Tail fins: Also called the tail fins, caudal fins are attached to the end of the caudal peduncle and used for propulsion. The caudal peduncle is the narrow part of the fish's body. The hypural joint is the joint between the caudal fin and the last of the vertebrae. The hypural is often fan-shaped. The tail may be heterocercal, reversed heterocercal, protocercal, diphycercal, or homocercal.
- Heterocercal: vertebrae extend into the upper lobe of the tail, making it longer (as in sharks)
- Reversed heterocercal: vertebrae extend into the lower lobe of the tail, making it longer (as in the Anaspida)
- Protocercal: vertebrae extend to the tip of the tail; the tail is symmetrical but not expanded (as in lancelets).
- Diphycercal: vertebrae extend to the tip of the tail; the tail is symmetrical and expanded (as in the bichir, lungfish, lamprey and Palaeozoic fishes had a diphycercal heterocercal tail.[28]
- Homocercal: vertebrae extend a very short distance into the upper lobe of the tail; tail still appears superficially symmetric. Most fish have a homocercal tail, but it can be expressed in a variety of shapes. The tail fin can be rounded at the end, truncated (almost vertical edge, as in salmon), forked (ending in two prongs), emarginate (with a slight inward curve), or continuous (dorsal, caudal, and anal fins attached, as in eels).
- Anal fins: Located on the ventral surface behind the anus, this fin is used to stabilize the fish while swimming.
- sea robins and flying gurnards.
- "Cephalic fins": The "horns" of manta rays and their relatives, sometimes called cephalic fins, are actually a modification of the anterior portion of the pectoral fin.
- Pelvic/Ventral fins: Found in pairs on each side ventrally below the pectoral fins, pelvic fins are homologous to the hindlimbs of tetrapods. They assist the fish in going up or down through the water, turning sharply, and stopping quickly. In gobies, the pelvic fins are often fused into a single sucker disk that can be used to attach to objects.
- characins and catfishes. Its function has remained a mystery, and is frequently clipped off to mark hatchery-raised fish, though data from 2005 showed that trout with their adipose fin removed have an 8% higher tailbeat frequency.[29] Additional research published in 2011 has suggested that the fin may be vital for the detection of and response to stimuli such as touch, sound and changes in pressure. Canadian researchers identified a neural network in the fin, indicating that it likely has a sensory function, but are still not sure exactly what the consequences of removing it are.[30]
- Caudal keel: A lateral ridge, usually composed of scutes, on the caudal peduncle just in front of the tail fin. Found on some types of fast-swimming fish, it provides stability and support to the caudal fin, much like the keel of a ship. There may be a single paired keel, one on each side, or two pairs above and below.
- sauries, they are rayless, non-retractable, and found between the last dorsal or anal fin and the caudal fin.
Internal organs
Intestines
As with other vertebrates, the
Pyloric caeca
Many fish have a number of small outpocketings, called pyloric caeca, along their intestine. The purpose of the caeca is to increase the overall surface area of the intestines, thereby increasing the absorption of nutrients.[32][33]
The number of pyloric caeca varies widely between species, and in some species of fish no caeca are present at all. Species with few or no caeca compensate for their lack by having longer intestines, or by have taller or more convoluted intestinal villi, thereby achieving similar levels of absorptive surface area.[32][33]
Lungfish also have a pouch located at the beginning of their intestine, which is also called a pyloric
Stomach
As with other vertebrates, the relative positions of the
Kidneys
The kidneys of fish are typically narrow, elongated organs, occupying a significant portion of the trunk. They are similar to the mesonephros of higher vertebrates (reptiles, birds, and mammals). The kidneys contain clusters of nephrons, serviced by collecting ducts which usually drain into a mesonephric duct. However, the situation is not always so simple. In cartilaginous fish, there is also a shorter duct which drains the posterior (metanephric) parts of the kidney, and joins with the mesonephric duct at the bladder or cloaca. Indeed, in many cartilaginous fish, the anterior portion of the kidney may degenerate or cease to function altogether in the adult.[35] Hagfish and lamprey kidneys are unusually simple. They consist of a row of nephrons, each emptying directly into the mesonephric duct.[35] Like the Nile tilapia, the kidney of some fish shows its three parts; head, trunk, and tail kidneys.[36] Fish do not have a discrete adrenal gland with distinct cortex and medulla, similar to those found in mammals. The interrenal and chromaffin cells are located within the head kidney.[36]
Urinary bladder
The gills of most teleost fish help to eliminate ammonia from the body, and fish live surrounded by water, but most still have a distinct bladder for storing waste fluid. The urinary bladder of teleosts is permeable to water, though this is less true for freshwater dwelling species than saltwater species.[37]: p. 219 In freshwater fish the bladder is a key site of absorption for many major ions[38] in marine fish urine is held in the bladder for extended periods to maximise water absorption.[38] The urinary bladders of fish and tetrapods are thought to be analogous while the former's swim-bladders and latter's lungs are considered homologous.
Most fish also have an organ called aSpleen
The
Liver
The liver is a large
Heart


Fish have what is often described as a two-chambered heart,[43] consisting of one atrium to receive blood and one ventricle to pump it,[44] in contrast to three chambers (two atria, one ventricle) of amphibian and most reptile hearts and four chambers (two atria, two ventricles) of mammal and bird hearts.[43] However, the fish heart has entry and exit compartments that may be called chambers, so it is also sometimes described as three-chambered,[44] or four-chambered,[45] depending on what is counted as a chamber. The atrium and ventricle are sometimes considered "true chambers", while the others are considered "accessory chambers".[46]
The four compartments are arranged sequentially:
- Atrium: A thicker-walled, muscular chamber that sends blood to the ventricle.[44]
- Ventricle: A thick-walled, muscular chamber that pumps the blood to the fourth part, the outflow tract.[44] The shape of the ventricle varies considerably, usually tubular in fish with elongated bodies, pyramidal with a triangular base in others, or sometimes sac-like in some marine fish.[45]
- Outflow tract (OFT): Goes to the ventral aorta and consists of the tubular conus arteriosus, bulbus arteriosus, or both.[45] The conus arteriosus, typically found in more primitive species of fish, contracts to assist blood flow to the aorta, while the bulbus anteriosus does not.[46][47]
Ostial valves, consisting of flap-like connective tissues, prevent blood from flowing backward through the compartments.
The ventral aorta delivers blood to the gills where it is oxygenated and flows, through the dorsal aorta, into the rest of the body. (In tetrapods, the ventral aorta is divided in two; one half forms the ascending aorta, while the other forms the pulmonary artery).[41]
The circulatory systems of all vertebrates are
In the adult fish, the four compartments are not arranged in a straight row, instead forming an S-shape with the latter two compartments lying above the former two. This relatively simpler pattern is found in cartilaginous fish and in the ray-finned fish. In teleosts, the conus arteriosus is very small and can more accurately be described as part of the aorta rather than of the heart proper. The conus arteriosus is not present in any amniotes, presumably having been absorbed into the ventricles over the course of evolution. Similarly, while the sinus venosus is present as a vestigial structure in some reptiles and birds, it is otherwise absorbed into the
Swim bladder
The swim bladder or gas bladder is an internal organ that contributes to the ability of a fish to control its
Weberian apparatus
Fishes of the
Reproductive organs

Fish reproductive organs include

Testes

Most male fish have two testes of similar size. In the case of sharks, the testis on the right side is usually larger. The primitive jawless fish have only a single testis located in the midline of the body, although even this forms from the fusion of paired structures in the embryo.[41]
Under a tough membranous shell, the
However, most fish do not possess seminiferous tubules. Instead, the sperm are produced in spherical structures called sperm ampullae. These are seasonal structures, releasing their contents during the breeding season and then being reabsorbed by the body. Before the next breeding season, new sperm ampullae begin to form and ripen. The ampullae are otherwise essentially identical to the seminiferous tubules in higher vertebrates, including the same range of cell types.[57]
In terms of
Ovaries
Many of the features found in ovaries are common to all vertebrates, including the presence of
Fish ovaries may be of three types: gymnovarian, secondary gymnovarian or cystovarian. In the first type, the
Nervous system

Central nervous system
Fish typically have quite small brains relative to body size compared with other vertebrates, typically one-fifteenth the brain mass of a similarly sized bird or mammal.
Fish brains are divided into several regions. At the front are the
.The forebrain is connected to the
The hindbrain or metencephalon is particularly involved in swimming and balance.[59] The cerebellum is a single-lobed structure that is typically the biggest part of the brain.[59] Hagfish and lampreys have relatively small cerebella, while the mormyrid cerebellum is massive and apparently involved in their electrical sense.[59]
The
Vertebrates are the only chordate group to exhibit a proper brain. A slight swelling of the anterior end of the
Cerebellum
The circuits in the cerebellum are similar across all classes of vertebrates, including fish, reptiles, birds, and mammals.[64] There is also an analogous brain structure in cephalopods with well-developed brains, such as octopuses.[65] This has been taken as evidence that the cerebellum performs functions important to all animal species with a brain.
There is considerable variation in the size and shape of the cerebellum in different vertebrate species. In amphibians, lampreys, and hagfish, the cerebellum is little developed; in the latter two groups, it is barely distinguishable from the brain-stem. Although the
The cerebellum of cartilaginous and bony fishes is extraordinarily large and complex. In at least one important respect, it differs in internal structure from the mammalian cerebellum: The fish cerebellum does not contain discrete
Most species of fish and amphibians possess a lateral line system that senses
Identified neurons
A neuron is "identified" if it has properties that distinguish it from every other neuron in the same animal—properties such as location, neurotransmitter, gene expression pattern, and connectivity—and if every individual organism belonging to the same species has one and only one neuron with the same set of properties.[67] In vertebrate nervous systems, very few neurons are "identified" in this sense (in humans, there are believed to be none). In simpler nervous systems, some or all neurons may be thus unique.[68]
In vertebrates, the best known identified neurons are the gigantic
Mauthner cells have been described as command neurons. A command neuron is a special type of identified neuron, defined as a neuron that is capable of driving a specific behavior all by itself.[70] Such neurons appear most commonly in the fast escape systems of various species—the squid giant axon and squid giant synapse, used for pioneering experiments in neurophysiology because of their enormous size, both participate in the fast escape circuit of the squid. The concept of a command neuron has, however, become controversial, because of studies showing that some neurons that initially appeared to fit the description were really only capable of evoking a response in a limited set of circumstances.[71]
Immune system
Immune organs vary by type of fish.
Cartilaginous fish (sharks and rays) have a more advanced immune system. They have three specialized organs that are unique to
Like chondrostean fish, the major immune tissues of bony fish (
See also
- Anatomical terms of location
- Decapod anatomy
- Digital Fish Library
- Evolution of fish
- Fish development
- Fish measurement
- Fish physiology
- Gastropod anatomy
- Ichthyology terms
- Panderichthys digits
- Shark anatomy
References
- from the original on 3 July 2021. Retrieved 3 July 2021.
- ^ ISBN 978-0-03-030504-7.
- ^ "The fish heart". ThinkQuest. Oracle. Archived from the original on 28 April 2012. Retrieved 27 June 2013.
- ^ ISBN 9788171338917. Archived from the originalon 22 April 2016.
- ISBN 9780520936966
- ^ Waggoner, Ben. "Vertebrates: Fossil Record". UCMP. Archived from the original on 29 June 2011. Retrieved 15 July 2011.
- ISBN 978-0-19-878555-2.
- ^ Waggoner, Ben. "Vertebrates: More on Morphology". UCMP. Archived from the original on 6 August 2012. Retrieved 13 July 2011.
- ^ a b c Romer, A.S. (1949): The Vertebrate Body. W.B. Saunders, Philadelphia. (2nd ed. 1955; 3rd ed. 1962; 4th ed. 1970)
- ISBN 978-0-03-022369-3.
- ^ ISBN 978-0-03-910284-5.
- S2CID 5613153.
- PMID 1496398.
- PMID 19741680.
- ^ ISBN 978-0-03-910284-5.
- (PDF) from the original on 27 September 2011. Retrieved 11 January 2011.
- N. A. Campbell and J. B. Reece(2005). Biology, Seventh Edition. Benjamin Cummings, San Francisco, California.
- PMID 18026166.
- ^ Helfman, Collette, Facey and Bowen, 2009, The Diversity of Fishes: Biology, Evolution, and Ecology pp. 84–87.
- ^ "integument - Arthropods | Britannica". www.britannica.com. Retrieved 6 May 2022.
- ^ ISBN 978-1-259-70091-0.
- S2CID 20223479.
- ^ "integument - Skin derivatives and appendages | Britannica". www.britannica.com. Retrieved 20 April 2022.
- ^ ISBN 978-0-03-910284-5.
- ^ "integument - Fishes | Britannica". www.britannica.com. Retrieved 20 April 2022.
- ISBN 978-1-259-70091-0.
- S2CID 205221027.
- ^ von Zittel KA, Woodward AS and Schloser M (1932) Text-book of Paleontology Volume 2, Macmillan and Company. Page 13.
- from the original on 26 July 2008. Retrieved 8 February 2011.
- ^ "Removal of trout, salmon fin touches a nerve". Archived from the original on 20 July 2011.
- ISBN 978-1-85233-241-9. Archivedfrom the original on 14 April 2021. Retrieved 9 January 2009.
- ^ ISBN 978-0-03-910284-5.
- ^ (PDF) from the original on 23 May 2018. Retrieved 22 May 2018.
- ISBN 978-0-03-910284-5.
- ^ ISBN 978-0-03-910284-5.
- ^ from the original on 26 April 2021. Retrieved 26 April 2021.
- ISBN 978-3-662-05014-9.
- ^ PMID 37457024.
- ^ Owen, Richard (1843). Lectures on the comparative anatomy and physiology of the invertebrate animals. London: Longman, Brown, Green, and Longmans. pp. 283–284.
- Internet Encyclopedia of Science
- ^ ISBN 978-0-03-910284-5.
- from the original on 3 July 2021. Retrieved 3 July 2021 – via Elsevier Science Direct.
- ^ ISBN 978-1-85996-325-8. Archivedfrom the original on 6 December 2016. Retrieved 13 March 2016.
- ^ ISBN 978-0-12-529650-2. Archivedfrom the original on 6 December 2016. Retrieved 13 March 2016.
- ^ ISBN 978-0-08-092323-9. Archivedfrom the original on 6 December 2016. Retrieved 13 March 2016.
- ^ ISBN 978-81-7133-800-9. Archivedfrom the original on 6 December 2016. Retrieved 13 March 2016.
- S2CID 9676359.
- ISBN 978-0-87893-249-8.
- ISBN 978-0-07-304058-5.
- S2CID 84010604.
- ^ ISBN 978-0-471-25031-9.)
{{cite book}}: CS1 maint: multiple names: authors list (link - ^ ISSN 0101-8175.
- ISSN 0044-8486.
- S2CID 235311767.
- PMID 22634430.
- PMID 22919339.
- ^ ISBN 978-0-03-910284-5.
- ISSN 0102-0935.
- ^ a b c d e f g h i j k Helfman, Collette & Facey 1997, pp. 48–49
- ^ Helfman, Collette & Facey 1997, p. 191
- ^ a b Hildebrand, M. & Gonslow, G. (2001): Analysis of Vertebrate Structure. 5th edition. John Wiley & Sons, Inc. New York City
- ^ "Keeping an eye on evolution". PhysOrg.com. 3 December 2007. Archived from the original on 15 March 2012. Retrieved 4 December 2007.
- ^ "Hyperotreti - Hagfishes". Archived from the original on 6 February 2013. Retrieved 14 December 2012.
- ^ PMID 18275284.
- S2CID 43112389.
- PMID 18537139.
- ISBN 978-0-306-31001-0.
- ^ "Wormbook: Specification of the nervous system". Archived from the original on 17 July 2011. Retrieved 14 December 2012.
- ISBN 978-0-262-69227-4.
- ^ Stein, p. 112
- ISBN 978-0-521-62726-9.
- ISBN 9780123504395.
- ^ D.P. Anderson. Fish Immunology. (S. F. Snieszko and H. R. Axelrod, eds), Hong Kong: TFH Publications, Inc. Ltd., 1977.
- .
- S2CID 7965762.
- S2CID 428224.
Works cited
- Helfman, G.; Collette, B.; Facey, D. (1997). The Diversity of Fishes (1st ed.). Wiley-Blackwell. ISBN 978-0-86542-256-8.
External links
- Mongabay.com Fish anatomy Mongabay
- Stunning Fish X-rays Smithsonian exhibit, LiveScience, 13 June 2011.
Anatomy and morphology | ||
|---|---|---|
| Fields |
|  |
| Bacteria and fungi | ||
| Protists | ||
| Plants |
| |
| Invertebrates |
| |
Mammals | ||
Other vertebrates |
| |
| Glossaries | ||
| Related topics | ||