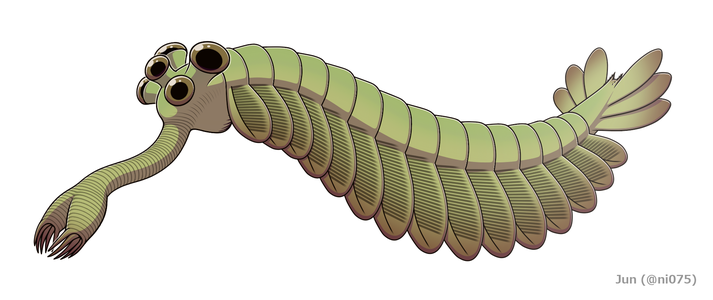
Middle Cambrian Burgess ShaleLagerstätte (505 million years ago) of British Columbia.[1]Opabinia was a soft-bodied animal, measuring up to 7 cm in body length, and its segmented trunk had flaps along the sides and a fan-shaped tail. The head shows unusual features: five eyes, a mouth under the head and facing backwards, and a clawed proboscis that probably passed food to the mouth. Opabinia probably lived on the seafloor, using the proboscis to seek out small, soft food.[2] Fewer than twenty good specimens have been described; 3 specimens of Opabinia are known from the Greater Phyllopod bed, where they constitute less than 0.1% of the community.[3]
When the first thorough examination of Opabinia in 1975 revealed its unusual features, it was thought to be unrelated to any known phylum,[4] or perhaps a relative of arthropod and annelid ancestors.[2] However, later studies since late 1990s consistently support its affinity as a member of basal arthropods, alongside the closely related radiodonts (Anomalocaris and relatives) and gilled lobopodians (Kerygmachela and Pambdelurion).[5][6][7][8][1][9][10]
In the 1970s, there was an ongoing debate about whether
stem groups, suggested that the Early Cambrian was a time of relatively fast evolution, but one that could be understood without assuming any unique evolutionary processes.[11]
History of discovery
In 1911,
paleontologists described specimens they found in the Norilsky region of Siberia and labelled Opabinia norilica,[14] but these fossils were poorly preserved, and Whittington did not feel they provided enough information to be classified as members of the genusOpabinia.[2]
Occurrence
All the recognized Opabinia specimens found so far come from the "Phyllopod bed" of the Burgess Shale, in the Canadian Rockies of British Columbia.[2]
In 1997, Briggs and Nedin reported from South AustraliaEmu Bay Shale a new specimen of Myoscolex that was much better preserved than previous specimens, leading them to conclude that it was a close relative of Opabinia[15]—although this interpretation was later questioned by Dzik, who instead concluded that Myoscolex was an annelid worm.[16]
Morphology
Restoration
Size estimation
Suggested proboscis mobility of Opabinia
Opabinia looked so strange that the audience at the first presentation of Whittington's analysis laughed.
digestive tract formed a U-bend on its way towards the rear of the animal. The proboscis appeared sufficiently long and flexible to reach the mouth.[2]
The main part of the body was typically about 5 millimetres (0.20 in) wide and had 15 segments, on each of which there were pairs of flaps (lobes) pointing downwards and outwards. The flaps overlapped so that the front of each was covered by the rear edge of the one ahead of it. The body ended with what looked like a single conical segment bearing three pairs of overlapping tail fan blades that pointed up and out, forming a tail like a V-shaped double fan.[2]
Various interpretations on the flap and gill structures of Opabinia regalis A: Whittington (1975),[2] B: Bergström (1986),[17] C: Budd (1996),[5] D: Zhang & Briggs (2007),[7] E: Budd & Daley (2011)[8]
Opabinia cross-section based on Budd and Daley (2011)[8]
Interpretations of other features of Opabinia fossils differ. Since the animals did not have mineralized armor nor even tough organic exoskeletons like those of other arthropods, their bodies were flattened as they were buried and fossilized, and smaller or internal features appear as markings within the outlines of the fossils.[2][5]
Whittington (1975) interpreted the gills as paired extensions attached dorsally to the bases of all but the first flaps on each side, and thought that these gills were flat underneath, had overlapping layers on top.
diverticula
individualized from the triangular extension.
= Body
= Base (basipod)
= Gill branch (exopod/exite)
// = Gill filaments
= Leg (endopod)
biramous
limb
Whittington (1975) found evidence of near-triangular features along the body, and concluded that they were internal structures, most likely sideways extensions of the gut (
biramous limbs seen in Marrella, trilobites, and crustaceans.[5] Zhang and Briggs (2007) analyzed the chemical composition of the "triangles", and concluded that they had the same composition as the gut, and therefore agreed with Whittington that they were part of the digestive system. Instead they regarded Opabinia's lobe+gill arrangement as an early form of the arthropod limbs before it split into a biramous structure.[7] However, this similar chemical composition is not only associated with the digestive tract; Budd and Daley (2011) suggest that it represents mineralization forming within fluid-filled cavities within the body, which is consistent with hollow lobopods as seen in unequivocal lobopodian fossils. They also clarify that the gut diverticula of Opabinia are series of circular gut glands individualized from the "triangles". While they agreed on the absence of terminal claws, the presence of lobopods in Opabinia remain as a plausible interpretation.[8]
Lifestyle
The way in which the
diverticula may increase the efficiency of food digestion and intake of nutrition.[22] Whittington (1975) believing that Opabinia had no legs, thought that it crawled on its lobes and that it could also have swum slowly by flapping the lobes, especially if it timed the movements to create a wave with the metachronal movement of its lobes.[2] On the other hand, he thought the body was not flexible enough to allow fish-like undulations of the whole body.[2]
Considering how paleontologists' reconstructions of Opabinia differ, it is not surprising that the animal's classification was highly debated during the 20th century.
Ecdysozoan, respectively) which were still thought to be close relatives (united under Articulata) at that time.[25]
tardigrades (water bears) may be Opabinia's closest living evolutionary relatives.[5] On the other hand, Hou et al. (1995, 2006) suggested Opabinia is a member of unusual cycloneuralian worms with convergent arthropod features.[29][18]
Although Zhang and Briggs (2007) disagreed with Budd's diagnosis that Opabinia's "triangles" were legs, the resemblance they saw between Opabinia's lobe+gill arrangement and arthropods'
biramous limbs led them to conclude that Opabinia was very closely related to arthropods. In fact they presented a family tree very similar to Budd's except that theirs did not mention tardigrades.[7] Regardless of the different morphological interpretations, all major restudies since 1980s similarly concluded that the resemblance between Opabinia and arthropods (e.g., stalked eyes, dorsal segmentation, posterior mouth, fused appendages, gill-like limb branches) are taxonomically significant.[17][5][7][8][1]
Since the 2010s, the suggested close relationship between Opabinia and tardigrades/cycloneuralians is no longer supported, while the affinity of Opabinia as a stem-group arthropod alongside Radiodonta (a clade that includes Anomalocaris and its relatives[30]) and gilled lobopodians is widely accepted,[1][9][10] as consistently shown by multiple phylogenetic analyses,[31][32][33][34][35][36][37][38][39][40][41][42] as well as new discoveries such as the presence of arthropod-like gut glands[8][22] and the intermediate taxon Kylinxia.[40]
In 2022, Paleontologists described a similar looking animal which was discovered in Cambrian-aged rocks of Utah.[43] The fossil was named Utaurora comosa, and was found within the Wheeler Shale.[43] The stem-arthropod was actually first described in 2008, but at the time it was originally considered a specimen of Anomalocaris.[44] This discovery could suggest there were other animals that looked like Opabinia, and its family may have been more diverse.[43]
taxa such as Marrella and Yohoia, cannot be accommodated in modern groups. This was one of the primary reasons why Gould in his book on the Burgess Shale, Wonderful Life, considered that Early Cambrian life was much more disparate and "experimental" than any later set of animals and that the Cambrian explosion was a truly dramatic event, possibly driven by unusual evolutionary mechanisms.[4] He regarded Opabinia as so important to understanding this phenomenon that he wanted to call his book Homage to Opabinia.[49]
However, other discoveries and analyses soon followed, revealing similar-looking animals such as Anomalocaris from the Burgess Shale and Kerygmachela from Sirius Passet.[17][27] Another Burgess Shale animal, Aysheaia, was considered very similar to modern Onychophora,[50] which are regarded as close relatives of arthropods.[51] Paleontologists defined a group called lobopodians to include fossil panarthropods that are thought to be close relatives of onychophorans, tardigrades and arthropods but lack jointed limbs. This group was later widely accepted as a paraphyletic grade that led to the origin of extant panarthropod phyla.
While this discussion about specific fossils such as Opabinia and Anomalocaris was going on in late 20 century, the concept of
stem groups was introduced to cover evolutionary "aunts" and "cousins". A crown group is a group of closely related living animals plus their last common ancestor plus all its descendants. A stem group contains offshoots from members of the lineage earlier than the last common ancestor of the crown group; it is a relative concept, for example tardigrades are living animals that form a crown group in their own right, but Budd (1996) regarded them also as being a stem group relative to the arthropods.[5][52] Viewing strange-looking organisms like Opabinia in this way makes it possible to see that, while the Cambrian explosion was unusual, it can be understood in terms of normal evolutionary processes.[11]
^Miroshnikov, L. D. and Krawzov, A. G. (1960). Rare paleontological remains and traces of life in late Cambrian deposits of the northwestern Siberian platform. Palaeontology and biostratigraphy of the Soviet Arctic, 3, pp. 28–41.
^Hutchinson, George Evelyn (1930). Restudy of some Burgess shale fossils. Proceedings of the United States National Museum78 (2854): 1–24. doi:10.5479/si.00963801.78-2854.1.
^Simonetta AM. 1970 Studies on non trilobite arthropods of the Burgess Shale (Middle Cambrian). Palaeontogr. Ital. 66, 35–45.
^Budd, G. E. (1997). "Stem Group Arthropods from the Lower Cambrian Sirius Passet Fauna of North Greenland". In Fortey, R. A.; Thomas, R. H. (eds.). Arthropod Relationships – Special Volume Series 55. Systematics Association.
. and Cloud, P. E. (1968). "Pre-metazoan evolution and the origins of the Metazoa.". In Drake, E. T. (ed.). Evolution and Environment. New Haven, Conn.: Yale University Press. pp. 1–72.
^Durham, J. W. (1971). "The fossil record and the origin of the Deuterostomata". Proceedings of the North American Paleontological Convention, Part H: 1104–1132. and Glaessner, M. F. (1972). "Precambrian palaeozoology". In Jones, J. B.; McGowran, B. (eds.). Stratigraphic Problems of the Later Precambrian and Early Cambrian. Vol. 1. University of Adelaide. pp. 43–52.
^ abCraske, A. J.; Jefferies, R. P. S. (1989). "A new mitrate from the Upper Ordovician of Norway, and a new approach to subdividing a plesion". Palaeontology. 32: 69–99.
Further reading
Bergström, J. (1987). "The Cambrian Opabinia and Anomalocaris". Lethaia. 20 (2): 187–188.


 Restoration
Restoration Size estimation
Size estimation


![Various interpretations on the flap and gill structures of Opabinia regalis A: Whittington (1975),[2] B: Bergström (1986),[17] C: Budd (1996),[5] D: Zhang & Briggs (2007),[7] E: Budd & Daley (2011)[8]](//upload.wikimedia.org/wikipedia/commons/thumb/3/34/20210809_Opabinia_regalis_flap_gill_interpretation.png/780px-20210809_Opabinia_regalis_flap_gill_interpretation.png)
![Opabinia cross-section based on Budd and Daley (2011)[8]](//upload.wikimedia.org/wikipedia/commons/thumb/3/30/20210807_Opabinia_regalis_trunk_cross_section.png/420px-20210807_Opabinia_regalis_trunk_cross_section.png) Opabinia cross-section based on Budd and Daley (2011)[8]
Opabinia cross-section based on Budd and Daley (2011)[8]![Various interpretations on the flap and gill structures of Opabinia regalis A: Whittington (1975),[2] B: Bergström (1986),[17] C: Budd (1996),[5] D: Zhang & Briggs (2007),[7] E: Budd & Daley (2011)[8]](/File:20210809_Opabinia_regalis_flap_gill_interpretation.png)
![Opabinia cross-section based on Budd and Daley (2011)[8]](/File:20210807_Opabinia_regalis_trunk_cross_section.png)







