
Radiodonta
| Radiodonta | |
|---|---|

| |
| Left to right, top to bottom: victoria | |
| Scientific classification | |
| Kingdom: | |
| Phylum: | |
| Class: | |
| Order: | †Radiodonta
Collins, 1996 |
| Families | |
| |
Radiodonta is an extinct
Etymology
The name Radiodonta (Latin for radius "spoke of a wheel" and Greek for odoús "tooth") refers to the radial arrangement of tooth plates (oral cone) surrounding the mouth,[6] although these features are suggested to be absent in some radiodont species.[4][1]
Definition
The original diagnosis of order Radiodonta in 1996 is as follows:[6]
Radiodontids are bilaterally symmetrical, elongate arthropods with a nonmineralized cuticle typically most robust in the jaws and claws. The body is subdivided into two
metameric, typically with about 13 segments laterally developing imbricating lobes for swimming and gills for respiration, and may end in a prominent three-part tail. Some forms have gnathobasic trunk limbs.
In 2014, the clade Radiodonta was defined
Description

Most radiodonts were significantly larger than the other
The body of a radiodont could be divided into two regions: head and trunk. The head is composed of only one body segment[17] known as the ocular somite, covered by sclerites (head carapace complex), bore arthropodized frontal appendages, ventral mouthparts (oral cone), and stalked compound eyes. The tapering trunk is composed of multiple body segments, each associated with pairs of flaps and gill-like structures (setal blades).[3]
Frontal appendage


The anterior structures on the head are a pair of frontal appendages which have been referred to as 'claws', 'grasping appendages', 'feeding appendages', or 'great appendages' in previous studies (the last term is discouraged since the homology between frontal appendages and the original, morphologically distinct
-
 Frontal appendages of Anomalocarididae, Amplectobeluidae, and possibly related species
Frontal appendages of Anomalocarididae, Amplectobeluidae, and possibly related species -
 Frontal appendages of Tamisiocarididae
Frontal appendages of Tamisiocarididae -
 Frontal appendages of Hurdiidae
Frontal appendages of Hurdiidae



Oral cone

The mouth is on the ventral side of the head, behind the attachment point of frontal appendages and is surrounded by a ring of tooth plates, forming the mouthpart known as oral cone ('jaws' in previous studies[6]). 3 or 4 tooth plates might be enlarged, giving the oral cone a triradial (e.g. Anomalocaris, Echidnacaris) or tetraradial (e.g. Hurdiidae, Lyrarapax) appearance.[23][12][24] The inner margin of tooth plates have spikes facing towards the mouth opening. Additional rows of internal tooth plates may occur in some hurdiid genera.[8][3] Detail reconstruction of some amplectobeluid oral cones are speculative, but they possibly did not present a typical radial arrangement.[4][1]
Head sclerites, eyes and trunk

Three head sclerite (carapace) complex formed by a central H-element (anterior sclerite or head shield) and a pair of P-elements (lateral sclerites) cover the dorsal and laterovental surface of the animal's head.[3] The P-elements may connect to each other as well as the H-element by a narrow anterior extension (P-element neck or 'beak').[8][3] The head sclerites are small and ovoid in Anomalocarididae and Amplectobeluidae,[4][3] but often enlarged in Hurdiidae, corresponded to their distinct body shapes (streamlined in Anomalocarididae/Amplectobeluidae but often compact in Hurdiidae).[3] The head bore two stalked compound eyes, which may have had mobility,[25] and are located between the gaps formed by the posterior regions of the H-element and P-elements.[8][3] The compound eyes of Echidnacaris are exceptionally unstalked.[13] Some species possess an additional median eye behind the H-element.[22]


Contrary to the original diagnosis, the division of body segments (segmental boundaries) can be visible externally
-
 Variations of radiodont body flaps
Variations of radiodont body flaps -
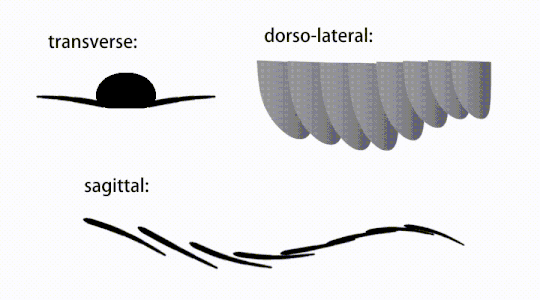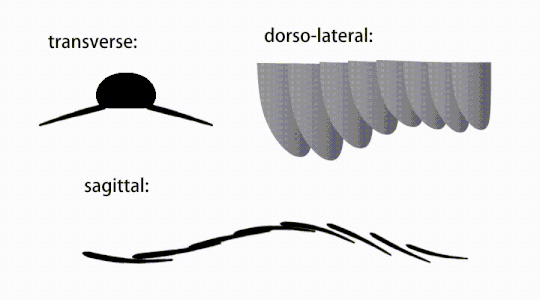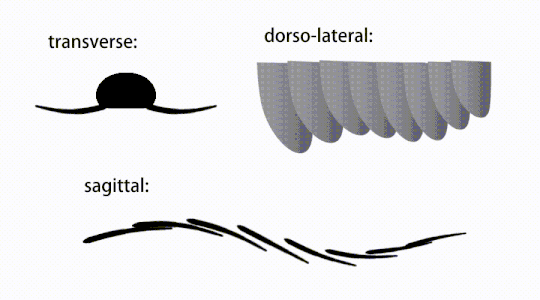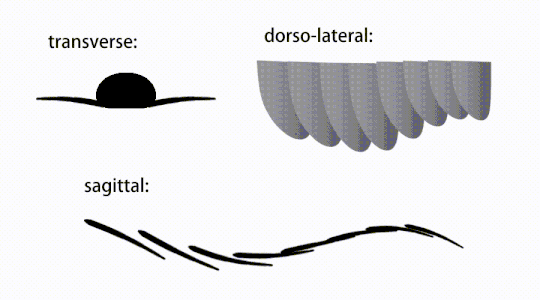 The movement of radiodont flapping appendages
The movement of radiodont flapping appendages -
 Ventral view of a generalized GLS-bearing radiodont, showing gnathobase‐like structures (GLSs) associated with reduced anterior flaps
Ventral view of a generalized GLS-bearing radiodont, showing gnathobase‐like structures (GLSs) associated with reduced anterior flaps



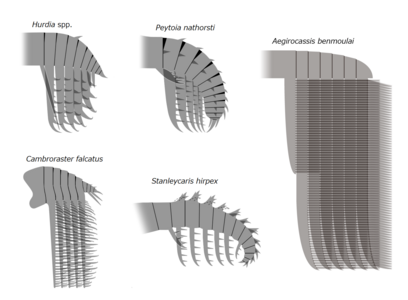
The trunk appendages were fin-like body flaps ('lateral flaps' or 'lobes' in some studies), usually one pair of ventral flaps per body segment, each slightly overlapping the one more anterior to it, but additional, non-overlapping sets of small dorsal flaps may occur in some Hurdiid species.[10] The flaps may have numerous vein-like structures (referred to as 'strengthening rays',[5] 'flap rays',[3] 'tranverse rods',[10] 'transverse lines'[28] or 'veins'[29]). The flaps on the neck region (referred to as 'reduced flaps',[4] 'neck flaps',[5] 'head flaps',[27] 'anterior flaps'[30] or 'differentiated flaps'[18]) are significantly reduced. In some species, jaw-like feeding appendages called gnathobase-like structures (GLSs) arose from each of the bases of their reduced neck flaps.[4][1] Numerous elongated blade-like extensions (referred to as lanceolate blades or lamellae[3]) arranged in a row, forming bands of gill-like structures known as setal blades, covered the dorsal surface of each body segment.[10] At least in Aegirocassis, each of the lanceolate blades are covered in wrinkles.[10] The ventral flaps may be homologous to the endopod of the biramous limbs of euarthropods and lobopodous limbs (lobopods) of gilled lobopodians, and the dorsal flaps and setal blades may be homologous to the exite and gill-bearing dorsal flaps of the former taxa.[31][10] The trunk may end either with a tail fan compose of 1 to 3 pairs of blades,[29][27][3] a pair of long furcae,[29][12][3] an elongated terminal structure,[27] or a featureless blunt tip.[10]
-
 Stanleycaris, Hurdia, Aegirocassis, Peytoia and Cambroraster are all examples of hurdiid radiodonts. These were the most diverse and long lasting of the radiodont families, surviving from the Cambrian up until the Devonian.
Stanleycaris, Hurdia, Aegirocassis, Peytoia and Cambroraster are all examples of hurdiid radiodonts. These were the most diverse and long lasting of the radiodont families, surviving from the Cambrian up until the Devonian. -
 Anomalocaris is a member of the anomalocarididae family, which at one point included all radiodonts, but now only includes a few genera such as Lenisicaris.
Anomalocaris is a member of the anomalocarididae family, which at one point included all radiodonts, but now only includes a few genera such as Lenisicaris. -
 Amplectobelua and Lyrarapax are representatives of the amplectobeluidae which is a very inclusive family of mainly Chinese radiodonts.
Amplectobelua and Lyrarapax are representatives of the amplectobeluidae which is a very inclusive family of mainly Chinese radiodonts. -
 Echidnacaris and Tamisiocaris are examples of the family tamisiocarididae which were exclusively suspension feeding radiodonts from the Cambrian.
Echidnacaris and Tamisiocaris are examples of the family tamisiocarididae which were exclusively suspension feeding radiodonts from the Cambrian.



Internal structures


Traces of
The
Paleoecology
Physiology

Radiodonts were interpreted as
Diet
Radiodonts had diverse feeding strategies, which could be categorized as
Oral cones of radiodonts may have been used for suction and/or biting.[23][38][3] Together with the great variety of frontal appendages in different species of radiodonts, differentiation of oral cones between species suggests preferences of different diets as well.[38][11] For example, the triradial oral cone of Anomalocaris with irregular, tuberculated toothplates and a small opening may have been adapted to small and nektonic prey,[23][11] while the rigid tetraradial oral cones of Peytoia, Titanokorys, Hurdia, and one isolated oral cone attributed to Cambroraster with a larger opening and sometimes additional tooth plates may have been capable to consume larger food items relative to their body size and probably benthic or endobenthic prey.[23][38][3]
Classification
Taxonomic affinities
| |||||||||||||||||||||||||||||||||||||||||||||
| Summarized phylogeny between Radiodonta and other Ecdysozoan taxa[42] |





-

-
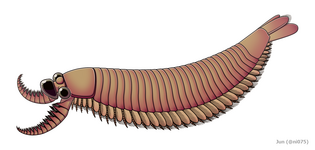 The radiodont/euarthropod intermediate Kylinxia, shares many of the characteristics found in both dinocaridids and euarthropods
The radiodont/euarthropod intermediate Kylinxia, shares many of the characteristics found in both dinocaridids and euarthropods -
 A life reconstruction of the basal deuteropod Erratus, which helped reveal the evolution of arthropod trunk appendages
A life reconstruction of the basal deuteropod Erratus, which helped reveal the evolution of arthropod trunk appendages -
 Middle Ordovicianthat shares traits with both opabiniids and radiodonts
Middle Ordovicianthat shares traits with both opabiniids and radiodonts



Most
-
 The siberiid lobopodians Siberion (upper left), Megadictyon (bottom center) and Jianshanopodia (upper right)
The siberiid lobopodians Siberion (upper left), Megadictyon (bottom center) and Jianshanopodia (upper right) -
-
 cheliceratepreviously thought to be radiodont's close relative
cheliceratepreviously thought to be radiodont's close relative



Taxa just basal to the radiodont, opabiniid and euarthropod branch are '
Previous studies may suggest radiodonts as a group other than stem-arthropods, such as a hitherto unknown
Interrelationships
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Phylogeny of Radiodonta after Moysiuk & Caron 2021[39] |





Traditionally, all radiodont species have been placed within one family,
The original description of the order Radiodonta included
-
 One of the poorly-known body parts (trunk appendage) of Cucumericrus decoratus; this species may not represent a true radiodont.
One of the poorly-known body parts (trunk appendage) of Cucumericrus decoratus; this species may not represent a true radiodont. -
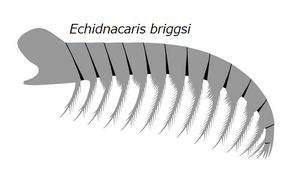 Frontal appendage ofEchidnacaris briggsi, a tamisiocaridradiodont that was once suggested to belong to the Anomalocaris genus until its description in 2023.
Frontal appendage ofEchidnacaris briggsi, a tamisiocaridradiodont that was once suggested to belong to the Anomalocaris genus until its description in 2023.

The first in-depth phylogenetic analysis of Radiodonta was conducted by Vinther et al. in 2014,
- Radiodonta
- ?Cucumericrus[26] (radiodont affinity questionable[65][64])
- ?Caryosyntrips[21] (radiodont affinity questionable[64])
- Anomalocarida
- Paranomalocaris[66] (placed within Anomalocarididae by some studies.[12][67])
- Laminacaris[68] (placed within Amplectobeluidae by some studies.[2])
- Houcaris (either placed within Anomalocarididae,[10][12][43] Amplectobeluidae[7][2] or Tamisiocarididae.[15][39])
- Innovatiocaris[69]
- Anomalocarididae
- Anomalocaris (in a broader sense, some species may placed within the other families.[7][10])
- Lenisicaris[20]
- Amplectobeluidae
- Lyrarapax[9] (position questioned by some studies.[1])
- Amplectobelua[26]
- Ramskoeldia[1] (position questioned by some studies.[3])
- Guanshancaris[67][70]
- Tamisiocarididae
- Peytoiidae[64]
- Aegirocassisinae[71]
- Peytoia
- Schinderhannes
- Hurdia
- Stanleycaris[72]
- Pahvantia[2]
- Ursulinacaris[19]
- Cambroraster[3]
- ?Zhenghecaris (putative hurdiid radiodont[65])
- Cordaticaris[73]
- Buccaspinea[74]
- Titanokorys[40]
| Species | Original description | Year named | Family | Age | Location | Frontal appendage | Head sclerite complex |
|---|---|---|---|---|---|---|---|
| Cucumericrus decoratus | Hou, Bergström, & Ahlberg | 1995[26] | (unassigned) | Cambrian Stage 3 | Unknown | Unknown | |
| Caryosyntrips serratus | Daley & Budd | 2010[21] | (unassigned) | Wuliuan–Drumian | 
|
Unknown | |
| Caryosyntrips camurus | Pates & Daley | 2017[41] | (unassigned) | Wuliuan | 
|
Incomplete[74] | |
| Caryosyntrips durus | Pates & Daley | 2017[41] | (unassigned) | Drumian | 
|
Unknown | |
| Paranomalocaris multisegmentalis | Wang, Huang, & Hu | 2013[66] | Anomalocarididae? | Cambrian Stage 4 | 
|
Unknown | |
| Paranomalocaris simplex | Jiao, Pates, Lerosey-Aubril, Ortega-Hernandez, Yang, Lan, Zhang | 2021[67] | Anomalocarididae? | Cambrian Stage 4 | 
|
Unknown | |
| Laminacaris chimera | Guo, Pates, Cong, Daley, Edgecombe, Chen, & Hou | 2018[68] | (controversial) | Cambrian Stage 3 | 
|
Unknown | |
| Innovatiocaris maotianshanensis | Zeng, Zhao, Zhu | 2022[69] | (unassigned) | Cambrian Stage 3 | 
|
P-element unknown[69] | |
| Innovatiocaris? multispiniformis | Zeng, Zhao, Zhu | 2022[69] | (unassigned) | Cambrian Stage 3 | 
|
Unknown | |
| Anomalocaris canadensis | Whiteaves | 1892[75] | Anomalocarididae | Wuliuan | 
|

| |
| Lenisicaris pennsylvanica (formerly Anomalocaris pennsylvanica)[20] | Resser | 1929 | Anomalocarididae | Cambrian Stage 3 | 
|
Unknown | |
| Lenisicaris lupata | Wu, Ma, Lin, Sun, Zhang, & Fu | 2021[20] | Anomalocarididae | Cambrian Stage 3 | 
|
Unknown | |
| Anomalocaris daleyae | Paterson, García-Bellidob & Edgecombe | 2023 | Anomalocarididae | Cambrian Stage 4 | 
|
Unknown | |
| Houcaris magnabasis (formerly Anomalocaris magnabasis)[15] | Pates, Daley, Edgecombe, Cong & Lieberman | 2019 | (controversial) | Cambrian Stage 4 | 
|
Unknown | |
| Houcaris saron (formerly Anomalocaris saron)[15] | Hou, Bergström, & Ahlberg | 1995 | (controversial) | Cambrian Stage 3 | 
|
Unknown | |
| Echidnacaris briggsi[24] | Nedin | 1995 | Tamisiocarididae | Cambrian Stage 4 | 
|
Possible H-element and unique lateral sclerites associated with compound eyes[13][24] | |
| Ramskoeldia platyacantha | Cong, Edgecombe, Daley, Guo, Pates, & Hou | 2018[1] | Amplectobeluidae | Cambrian Stage 3 | 
|
Incomplete[1] | |
| Ramskoeldia consimilis | Cong, Edgecombe, Daley, Guo, Pates, & Hou | 2018[1] | Amplectobeluidae | Cambrian Stage 3 | 
|
Incomplete[1] | |
| Lyrarapax unguispinus | Cong, Ma, Hou, Edgecombe, & Strausfield | 2014[9] | Amplectobeluidae | Cambrian Stage 3 | 
|
P-element neck unknown | |
| Lyrarapax trilobus | Cong, Daley, Edgecombe, Hou, & Chen | 2016[5] | Amplectobeluidae | Cambrian Stage 3 | 
|
P-element unknown | |
| Amplectobelua symbrachiata | Hou, Bergström, & Ahlberg | 1995[26] | Amplectobeluidae | Cambrian Stage 3 | 
|

| |
| Amplectobelua stephenensis | Daley & Budd | 2010[21] | Amplectobeluidae | Wuliuan | 
|
Unknown | |
| Guanshancaris kunmingensis | Zhang et al. | 2023[70] | Amplectobeluidae | Cambrian Stage 4 | 
|
Unknown | |
| Tamisiocaris borealis | Daley & Peel | 2010 | Tamisiocarididae | Cambrian Stage 3 | 
|
Incomplete[7] | |
| Ursulinacaris grallae | Pates, Daley & Butterfield | 2019 | Hurdiidae | Wuliuan | 
|
Unknown | |
| Schinderhannes bartelsi | Kühl, Briggs, & Rust | 2009[35] | Hurdiidae | Emsian | Incomplete[3] | Incomplete[3] | |
| Stanleycaris hirpex | Pates, Daley, & Ortega-Hernández | 2018[72] | Hurdiidae | Wuliuan | 
|
P-element is unknown, possibly absent[22] | |
Peytoia nathorsti
|
Walcott | 1911[76] | Hurdiidae | Wuliuan–Drumian | 
|
Incomplete[3] | |
| Peytoia infercambriensis (formerly Cassubia infercambriensis)[77] | Lendzion | 1975 | Hurdiidae | Cambrian Stage 3 | 
|
Unknown | |
| Aegirocassis benmoulai | Van Roy, Daley, & Briggs | 2015[10] | Hurdiidae (Aegirocassisinae) | Tremadocian | 
|

| |
| Hurdia victoria | Walcott | 1912[78] | Hurdiidae | Wuliuan–Drumian | 
|

| |
| Hurdia triangulata | Walcott | 1912[78] | Hurdiidae | Wuliuan |
|

| |
| Cambroraster falcatus | Moysiuk & Caron | 2019[3] | Hurdiidae | Wuliuan | 
|

| |
| Pahvantia hastata | Robison & Richards | 1981 | Hurdiidae | Drumian | 
|

| |
| Cordaticaris striatus | Sun, Zeng, & Zhao | 2020[73] | Hurdiidae | Drumian | Incomplete[73] | 
| |
| Zhenghecaris shankouensis | Vanner, Chen, Huang, Charbonnier, & Wang | 2006 | Hurdiidae | Cambrian Stage 3 | Unknown |  | |
| Buccaspinea cooperi | Pates, Lerosey-Aubril, Daley, Kier, Bonino & Ortega-Hernández | 2021[74] | Hurdiidae | Drumian | 
|
Unknown | |
| Titanokorys gainesi | Caron & Moysiuk | 2021[40] | Hurdiidae | Wuliuan | 
|

| |
| Pseudoangustidontus duplospineus | Van Roy & Tetlie | 2006 | Hurdiidae (Aegirocassisinae) | Tremadocian | 
|
Unknown | |
| Pseudoangustidontus izdigua | Potin, Gueriau & Daley | 2023 | Hurdiidae (Aegirocassisinae) | Tremadocian | 
|
Incomplete[71] |
History
-
 Frontal appendage ofAnomalocaris canadensis
Frontal appendage ofAnomalocaris canadensis -
 Oral cone ofPeytoia nathorsti
Oral cone ofPeytoia nathorsti -
 H-element of Hurdia victoria
H-element of Hurdia victoria -
![Paired frontal appendages from an unnamed hurdiid radiodont[39]](//upload.wikimedia.org/wikipedia/commons/thumb/1/1d/USNM_PAL_57490.jpg/267px-USNM_PAL_57490.jpg)



The history of radiodonts is complex. Incomplete specimens pertaining to different body parts of the same species had historically been interpreted as belonging to different species and even different phyla.
The first known radiodont specimens were collected from the
The Geological Survey of Canada initiated a revision of Burgess Shale fossils in 1966, overseen by
The taxon Radiodonta itself was coined in 1996 by Desmond Collins, after it was established that Anomalocaris and its kin represented a distinctive lineage with arthropod affinities rather than a hitherto unknown phylum.
Until the 2010s, radiodonts were typically considered to be uniformly large apex predators, but discoveries of new species over the course of that decade led to a considerable increase in the known ecological and morphological diversity of the group.[7][10][2][3][84][74][39][40]
References
- ^ S2CID 90258934.
- ^ PMID 30218075.
- ^ PMID 31362637.
- ^ PMID 28854872.
- ^ S2CID 88742430.
- ^ S2CID 131622496.
- ^ S2CID 205237459.
- ^ S2CID 206517995.
- ^ S2CID 4451239.
- ^ S2CID 205242881.
- ^ PMID 34284622.
- ^ ISSN 2095-5138.
- ^ PMID 32742697.
- PMC 10833464.
- ^ S2CID 235221043.
- PMID 34527273.
- ^ PMID 27989966.
- ^ S2CID 226248177.
- ^ PMID 31210962.
- ^ S2CID 233565727.
- ^ ISSN 1475-4983.
- ^ S2CID 250361698.
- ^ S2CID 2042726.
- ^ S2CID 259719252.
- ^ PMID 26276096.
- ^ ISSN 1103-5897.
- ^ S2CID 86683798.
- S2CID 128955512.
- ^ S2CID 1913482.
- ^ PMID 31914921.
- ^ Van Roy, Peter; Daley, Allison C.; Briggs, Derek E. G. (2013). Anomalocaridids had two sets of lateral flaps. 57th Annual Meeting of The Paleontological Association. Zurich, Switzerland.
- ^ PMID 24785191.
- ^ .
- PMID 16002096.
- ^ S2CID 47555807.
- PMID 29697774.
- PMID 26431634.
- ^ a b c d De Vivo, Giacinto; Lautenschlager, Stephan; Vinther, Jakob (2016-12-16). "Reconstructing anomalocaridid feeding appendage dexterity sheds light on radiodontan ecology".
- ^ S2CID 236552819.
- ^ PMID 34527273.
- ^ S2CID 135026011.
- ^ S2CID 7751936.
- ^ PMID 35135344.
- PMID 19854297.
- PMID 23055069.
- ^ S2CID 86725173.
- PMID 24077329.
- ISSN 1502-3931.
- S2CID 246779634.
- ^ S2CID 246608509.
- PMID 25959966.
- ^ S2CID 189926344.
- ^ S2CID 225478171.
- ^ CiteSeerX 10.1.1.693.5869
- S2CID 85645934.
- S2CID 55477207.
- PMID 19854297.
- PMID 29784780.
- ISSN 1103-5897.
- S2CID 82841481.
- S2CID 205244325.
- S2CID 4456458.
- PMID 31822253.
- ^ ISSN 0022-3360.
- ^ S2CID 133549817.
- ^ S2CID 83910779.
- ^ S2CID 236683029.
- ^ ISSN 2056-2799.
- ^ S2CID 252147346.
- ^ PMID 37106783.
- ^ ISSN 2296-701X.
- ^ a b Pates, Stephen; Daley, Allison; Ortega-Hernández, Javier (2018-03-03). Reply to Comment on "Aysheaia prolata from the Utah Wheeler Formation (Drumian, Cambrian) is a frontal appendage of the radiodontan Stanleycaris" with the formal description of Stanleycaris.
- ^ S2CID 224868404.
- ^ PMID 33552709.
- ^ a b c d Whiteaves, J. F. (1892). "Description of a new genus and species of phyllocarid Crustacea from the Middle Cambrian of Mount Stephen, B. C.". The Canadian Record of Science. 5 (4).
- ^ a b c d Walcott, C. D. 1911a. Middle Cambrian holothurians and medusae. Cambrian geoogy and paleontology II. Smithsonian Miscellaneous Collections, 57: 41-68.
- S2CID 130745134.
- ^ a b c WALCOTT, C. D. 1912. Middle Cambrian Branchiopoda, Malacostraca, Trilobita and Merostomata. Smithsonian Miscellaneous Collections, 57: 145–228.
- ^ ROLFE, W. D. I. 1962. Two new arthropod carapaces from the Burgess Shale (Middle Cambrian) of Canada. Breviora Museum of Comparative Zoology, 60: 1–9.
- JSTOR 1303799.
- ^ Briggs, D. E. G. (1979). "Anomalocaris, the largest known Cambrian arthropod". Palaeontology. 22 (3): 631–664.
- S2CID 86465719.
- S2CID 134299859.
- PMID 33268353.