
Stramenopile
| Stramenopiles Temporal range: Late Mesoproterozoic-present,
| |
|---|---|
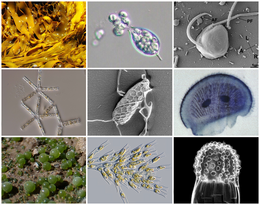 | |
| Diversity of stramenopiles | |
| Scientific classification | |
| Domain: | Eukaryota |
| Clade: | Diaphoretickes |
| Clade: | SAR |
| Clade: | Stramenopiles Patterson 1989[2] emend. Adl et al. 2005[3] |
| Phyla and subphyla[4] | |
| |
| Diversity | |
| >100000 species[1] | |
| Synonyms | |
The Stramenopiles, also called Heterokonts, are a
Stramenopiles are
Many stramenopiles are unicellular
The term 'heterokont' is used both as an adjective – indicating that a cell has two dissimilar flagella, and as the name of a taxon. The groups included in that taxon have however varied widely, creating the 'heterokont problem', now resolved by the definition of the stramenopiles.
History
The term 'stramenopile' was introduced by
The heterokont problem
The term 'heterokont' is used as both an adjective – indicating that a cell has two dissimilar flagella – and as the name of a taxon. The taxon 'Heterokontae' was introduced in 1899 by Alexander Luther for algae that are now considered the
Over time, the scope of application has changed, especially when in the 1970s ultrastructural studies revealed greater diversity among the algae with chromoplasts (chlorophylls a and c) than had previously been recognized. At the same time, a protistological perspective was replacing the 19th century one based on the division of unicellular eukaryotes into animals and plants. One consequence was that an array of heterotrophic organisms, many not previously considered as 'heterokonts', were seen as related to the 'core heterokonts' (those having anterior flagella with stiff hairs). Newly recognized relatives included the parasitic
Characteristics

The presumed
Most stramenopiles have two flagella near the apex.[22] They are usually supported by four microtubule roots in a distinctive pattern. There is a transitional helix inside the flagellum where the beating axoneme with its distinctive geometric pattern of nine peripheral couplets around two central microtubules changes into the nine-triplet structure of the basal body.[23]
Plastids
Many stramenopiles have
Ecology
Some stramenopiles are significant as autotrophs and as heterotrophs in natural ecosystems; others are parasitic. Blastocystis is a gastrointestinal parasite of humans;[27] opalines and proteromonads live in the intestines of cold-blooded vertebrates and have been described as parasitic;[28] oomycetes include some significant plant pathogens such as the cause of potato blight, Phytophthora infestans.[29] Diatoms are major contributors to global carbon cycles because they are the most important autotrophs in most marine habitats.[30] The brown algae, including familiar seaweeds like wrack and kelp, are major autotrophs of the intertidal and subtidal marine habitats.[31] Some of the bacterivorous stramenopiles, such as Cafeteria, are common and widespread consumers of bacteria, and thus play a major role in recycling carbon and nutrients within microbial food webs.[32][33]
Evolution
External
Stramenopiles are most closely related to Alveolates and Rhizaria, all of which have tubular mitochondrial cristae and collectively form the SAR supergroup, whose name is formed from their initials.[34][26][35] The ancestor of the SAR supergroup appears to have captured a unicellular photosynthetic red alga, and many Stramenopiles, as well as members of other SAR groups such as the Rhizaria, still have plastids which retain the double membrane of the red alga and a double membrane surrounding it, for a total of four membranes.[36] In addition, species of Telonemia, the sister group to SAR, exhibit heterokont flagella with tripartite mastigonemes, implying a more ancient origin of stramenopile characteristics.[37]
.jpg)
Internal
The following
| Stramenopiles |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Classification

The classification of the Stramenopiles according to Adl et al. (2019), with additions from newer research:[43] [4]
- Platysulcea Cavalier-Smith 2017[39]
- Bigyra Cavalier-Smith 1998, emend. 2006
- Opalozoa Cavalier Smith 1991, emend. 2006
- NanomonadeaCavalier-Smith 2012
- Opalinata Wenyon 1926, emend. Cavalier-Smith 1997 [=Slopalinida Patterson 1985]
- Bicosoecida Grasse 1926, emend. Karpov 1998
- Sagenista Cavalier-Smith 1995
- Labyrinthulomycetes Dick 2001
- Pseudophyllomitidae Shiratori et al. 2016[40]
- Opalozoa Cavalier Smith 1991, emend. 2006
- Gyrista Cavalier-Smith 1998
- Bigyromonada Cavalier-Smith 1998
- DevelopeaKarpov & Aleoshin 2016
- PirsonialesCavalier-Smith 1998, emend. 2006
- Pseudofungi Cavalier-Smith 1986
- HyphochytrialesSparrow 1960
- PeronosporomycetesDick 2001 [=Oomycetes Winter 1897, emend. Dick 1976]
- ActinophryidaeClaus 1874, emend. Hartmann 1926
- OchrophytaCavalier-Smith 1986, emend. Cavalier-Smith & Chao 1996
- ChrysistaCavalier-Smith 1986
- Chrysoparadoxophyceae Wetherbee 2019[44]
- ChrysophyceaePascher 1914
- Chloromorophyceae (nomen dubium)[45]
- EustigmatophyceaeHibberd & Leedale 1971
- PhaeophyceaeHansgirg 1886
- Phaeosacciophyceae R.A.Andersen, L.Graf & H.S.Yoon 2020[48]
- PhaeothamniophyceaeAndersen & Bailey in Bailey et al. 1998
- RaphidophyceaeChadefaud 1950, emend. Silva 1980
- Schizocladiophyceae Henry, Okuda & Kawai, 2003
- SynchromophyceaeS.Horn & C.Wilhelm 2007 [=Picophagea Cavalier-Smith 2006, emend. 2017]
- XanthophyceaeAllorge 1930, emend. Fritsch 1935 [Heterokontae Luther 1899; Heteromonadea Leedale 1983; Xanthophyta Hibberd 1990]
- DiatomistaDerelle et al. 2016, emend. Cavalier-Smith 2017
- Bolidophyceae Guillou et al. 1999
- DiatomeaeDumortier 1821 [=Bacillariophyta Haeckel 1878]
- Dictyochophyceae Silva 1980
- PelagophyceaeAndersen & Saunders 1993
- PinguiophyceaeKawachi et al. 2003
- Bigyromonada Cavalier-Smith 1998
Notes
- land plants.
References
- ^ . Retrieved 2 March 2024.
- ISBN 978-0198577133.
- Wikidata Q22065654.
- ^ PMID 28875267.
- S2CID 221852241.
- ^ David, J.C. (2002). "A preliminary catalogue of the names of fungi above the rank of order". Constancea. 83: 1–30.
- ISBN 978-0-948737-00-8.
- ISBN 978-0-521-30419-1.
- ISBN 978-0471522294.
- ISBN 978-94-015-9733-3.
- ^ "Stramenipila M.W. Dick (2001)". MycoBank. International Mycological Association.
- ^ Patterson, D. J. (1989). "Stramenopiles: chromophytes from a protistological perspective". In Green, J. C.; Leadbeater, B. S. C.; Diver, W. L. (eds.). The chromophyte algae: problems and perspectives. Oxford: Clarendon Press. pp. 357–379.
- S2CID 4367158.
- ^ David, J. C. (2002). "A preliminary catalogue of the names of fungi above the rank of order". Constancea (83): 1–30.
- ^ Luther, Alexander F. (1899). Über Chlorosaccus eine neue Gattung der Süsswasseralgen nebst einiger Bemerkungen zur Systematik verwandter Algen [About Chlorosaccus a new genus of freshwater algae together with some comments on the systematics of related algae] (in German). Stockholm: Norstedt. pp. 1–22.
- ^ Copeland, H. F. (1956). The Classification of Lower Organisms. Palo Alto, California: Pacific Books.
- ^ Blackwell, W. H. (2009). "Chromista revisited: A dilemma of overlapping putative kingdoms, and the attempted application of the botanical code of nomenclature" (PDF). Phytologia. 91 (2).
- ISBN 978-0-1239-1250-3.
- S2CID 52947780.
- PMID 5123323.
- PMID 20663714.
- ISBN 9780123739445.
- S2CID 7936118.
- .
- PMID 28889907.
- ^ PMID 27512113.
- PMID 24883113.
- OCLC 13123309.
- PMID 30731850.
- S2CID 16849373.
- PMID 21820616.
- ^ Guiry, Wendy (7 April 2011). "Cafeteria T.Fenchel & D.J.Patterson 1988". AlgaeBase. Retrieved 17 March 2023.
- ^ Fenchel, T.; Patterson, D. J. (1988). "Cafeteria roenbergensis nov. gen., nov. sp., a heterotrophic microflagellate from marine plankton". Marine Microbial Food Webs. 3: 9–19.
- ^ Krylov, M. V.; Dobrovolskii, A. A.; Issi, I. V.; Michaelevich, B. I.; Podlipaev, S. A.; Reshetnyak, V. V.; Seravin, L. N.; et al. 1980. New concepts for the system of unicellular organisms. Trudy Zoologischkei Institut Akademiya Nayuk, SSSR 94:122–132.
- PMID 18522922.
- PMID 24016529.
- PMID 35291881.
- ^ PMID 23219323.
- ^ S2CID 202865459.
- ^ PMID 28822908.
- ^ Wikidata Q123562228.
- Wikidata Q124719861.
- PMID 30257078.
- S2CID 54477112.
- ^ Medlin LK, Desdevises Y (2018). "Sequence analysis confirms a new algal class". Vie et Milieu/Life & Environment.
- S2CID 236175098.
- S2CID 232101186.
- S2CID 227315556.