
Lysenin
Lysenin is a
Monomer
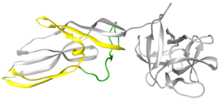
Lysenin is a
Membrane receptors
The natural
The region of the lysenin pore β-barrel expected to be immersed in the hydrophobic region of the membrane is the 'detergent belt', the 3.2 nm high region occupied by detergent in Cryogenic Electron Microscopy (Cryo-EM) studies of the pore.[10] On the other hand, sphingomyelin/Cholesterol bilayers are about 4.5 nm height.[11] This difference in height between the detergent belt and the sphingomyelin/cholesterol bilayer implies a bend of the membrane in the region surrounding the pore, called negative mismatch.[12] This bending results in a net attraction between pores that induce pores aggregation.
Binding, oligomerization and insertion

Membrane binding is a requisite to initiate PFT oligomerization. Lysenin monomers bind specifically to sphingomyelin via the receptor binding domain.[13] The final lysenin oligomer is constituted by nine monomers without quantified deviations.[14] When lysenin monomers bind to sphingomyelin-enriched membrane regions, they provide a stable platform with a high lateral mobility, hence favouring the oligomerization.[15] As with most PFTs, lysenin oligomerization occurs in a two-step process, as was recently imaged.
The process begins with monomers being adsorbed into the membrane by specific interactions, resulting in an increased concentration of monomers. This increase is promoted by the small area where the membrane receptor accumulates owing to the fact that the majority of PFT membrane receptors are associated with lipid rafts.[16] Another side effect, aside from the increase of monomer concentration, is the monomer-monomer interaction. This interaction increases lysenin oligomerization. After a critical threshold concentration is reached, several oligomers are formed simultaneously, although sometimes these are incomplete.[17] In contrast to PFTs of the cholesterol-dependent cytolysin family,[18] the transition from incomplete lysenin oligomers to complete oligomers has not been observed.
A complete oligomerization results in the so-called prepore state, a structure on the membrane. Determining the prepore's structure by X-ray or Cryo-EM is a challenging process that so far has not produced any results. The only available information about the prepore structure was provided by Atomic Force Microscopy (AFM). The measured prepore height was 90 Å; and the width 118 Å, with an inner pore of 50 Å.[17] A model of the prepore was built aligning the monomer structure (PDB: 3ZXD) with the pore structure (PDB: 5GAQ) by their receptor-binding domains (residues 160 to 297). A recent study in aerolysin suggests that the currently accepted model for the lysenin prepore should be revisited, according to the new available data on the aerolysin insertion.[19]
A
Insertion consequences
The ultimate consequences of lysenin pore formation are not well documented; however, it is thought to induce apoptosis via three possible hypotheses:
- Breaking the sphingomyelin asymmetry between the two leaflets of the lipid bilayer by punching holes in the membranelipid flip-flop (reorientation of a lipid from one leaflet of a membrane bilayer to the other).[28]
- Increasing the calcium concentration in the cytoplasm.[29]
- Decreasing the potassium concentration in the cytoplasm.[30]
Biological role
The biological role of lysenin remains unknown. It has been suggested that lysenin may play a role as a
Applications
Lysenin's conductive properties have been studied for years.[38] Like most pore-forming toxins, lysenin forms a non-specific channel that is permeable to ions, small molecules, and small peptides.[39] There have also been over three decades of studies into finding suitable pores for converting into nanopore sequencing systems that can have their conductive properties tuned by point mutation.[40] Owing to its binding affinity for sphingomyelin, lysenin (or just the receptor binding domain) has been used as a fluorescence marker to detect the sphingomyelin domain in membranes.[41]
References
![]() This article was submitted to WikiJournal of Science for external academic peer review in 2019 (reviewer reports). The updated content was reintegrated into the Wikipedia page under a CC-BY-SA-3.0 license (2019). The version of record as reviewed is:
Ignacio L. B. Munguira; et al. (17 August 2019). "Lysenin" (PDF). WikiJournal of Science. 2 (1): 6.
This article was submitted to WikiJournal of Science for external academic peer review in 2019 (reviewer reports). The updated content was reintegrated into the Wikipedia page under a CC-BY-SA-3.0 license (2019). The version of record as reviewed is:
Ignacio L. B. Munguira; et al. (17 August 2019). "Lysenin" (PDF). WikiJournal of Science. 2 (1): 6. {{cite journal}}: CS1 maint: unflagged free DOI (link
- PMID 16386304.
- PMID 28751253.
- .
- ^ PMID 22819216.
- PMID 27048994.
- ^ S2CID 22016219.
- S2CID 1866391.
- S2CID 16305100.
- PMID 26222645.
- PMID 27048994.
- PMID 23863113.
- PMID 26724385.
- PMID 22819216.
- S2CID 206699095.
- S2CID 22016219.
- S2CID 26547616.
- ^ PMID 24047991.
- PMID 26302195.
- PMID 27405240.
- ^ PMID 27048994.
- PMID 29117526.
- doi:10.1101/771725.
- PMID 26695013.
- ^ Munguira, I. L. B. (2017). Effect of Crowdedness in the Life Cycle of Lysenin Studied by High-Speed Atomic Force Microscopy (PhD). Aix-Marseille Universite.
- S2CID 208620873.
- S2CID 218539213.
- PMID 10893276.
- S2CID 16305100.
- S2CID 19079491.
- S2CID 13893235.
- ISBN 9780128032527.
- PMID 10684578.
- PMID 23965430.
- PMID 15261736.
- S2CID 19895826.
- S2CID 41192657.
- ISBN 978-0-412-56160-3.
- PMID 30126104.
- PMID 28550293.
- PMID 27153285.
- S2CID 1558393.