
Angomonas deanei
| Angomonas deanei | |
|---|---|
| |
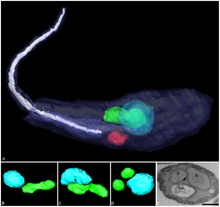
| Three-dimensional reconstruction of Angomonas deanei containing a bacterial endosymbiont (green) near its nucleus (blue). | |
| Scientific classification | |
| Domain: | Eukaryota |
| Phylum: | Euglenozoa |
| Class: | Kinetoplastea |
| Order: | Trypanosomatida |
| Family: | Trypanosomatidae |
| Genus: | Angomonas |
| Species: | A. deanei
|
| Binomial name | |
| Angomonas deanei (Carvalho, 1973)
Teixeira & Camargo, 2011[1] | |
| Synonyms | |
|
Crithidia deanei Carvalho, 1973 | |
Angomonas deanei is a
The species was first described as Crithidia deanei in 1973 by a Brazilian parasitologist Aurora L. M. Carvalho. A
Discovery
Angomonas deanei was originally described as Crithidia deanei. In 1973, a Brazilian graduate student Aurora Luiza de Moura Carvalho at the
As more structural and molecular details were studied, the distinction of A. deanei from other Crithidia species became more pronounced. In 1991, Maria Auxiliadora de Sousa and Suzana Corte-Real at the
Structure

The body of Angomonas deanei is elliptical in shape, with a prominent tail-like
While the protozoan has its separate mitochondria that provide
When the bacterium is killed using
Parasitism
Angomonas deanei was originally discovered from the digestive tract of the bug
Reproduction
The cellular reproduction shows a strong synergistic adaptation between the bacterium and the protozoan. The bacterium divides first, followed by the protozoan organelles, and lastly the nucleus. As a result the daughter protozoans contains exactly the same copies of the organelles and the bacterial endosymbiont.[36] The entire reproduction takes about 6 hours in an ideal culture medium; thus, a single protozoan is able to produce 256 daughter cells in a day, though it can differ slightly under its natural habitat.[21]
The endosymbiont and evolution
Symbiotic bacteria in the trypanosomatid protozoa are descended from a β-proteobacterium.
The symbiotic bacterium belongs to β-proteobacterium family
References
- ^ PMID 21420905.
- ^ Labinfo. "Angomonas deanei". labinfo.lncc.br. National Laboratory of Scientific Computation of the Ministry of Science and Technology, Brazil. Archived from the original on 2013-07-30. Retrieved 2013-07-08.
- PMID 29131838.
- PMID 23560078.
- PMID 29511084.
- PMID 26980834.
- S2CID 201731819.
- ^ PMID 23345457.
- ^ PMID 9127737.
- ^ PMID 16216441.
- ^ PMID 4594242.
- ^ Carvalho, Aurora Luiza de Moura (1973). "Estudos sobre a posição sistemática, a biologia e a transmissào de tripanosomatídeos encontrados em Zelus leucogrammus (Perty, 1834) (Hemiptera, Reduviidae)". Rev. Pat. Trop. (in Portuguese). 2 (2): 223–274.
- PMID 4278787.
- .
- S2CID 4210642.
- ^ Sousa, M.A. (1991). "Postnuclear kinetoplast in choanomastigotes of Crithidia deanei Carvalho, 1973. Proposal of a new genus". Revista do Instituto de Medicina Tropical de São Paulo. 33: S8.
- PMID 9057697.
- PMID 21757490.
- PMID 16436059.
- PMID 15755914.
- ^ PMID 27835948.
- PMID 33561222.
- PMID 24015778.
- PMID 24260300.
- PMID 21853145.
- PMID 23560078.
- PMID 25903782.
- PMID 17945218.
- PMID 27706380.
- PMID 10220875.
- PMID 16216441.
- PMID 29467742.
- S2CID 5830694.
- PMID 19070618.
- .
- PMID 20865129.
- PMID 8188611.
- PMID 16546131.
- PMID 7521530.
- ^ "GTDB - Tree at g__Kinetoplastibacterium". gtdb.ecogenomic.org.