
Holozoa
| Holozoans Temporal range:
| |
|---|---|
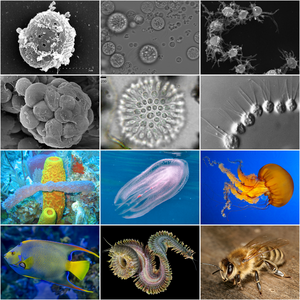 | |
| Holozoan diversity. Top half displays representatives of holozoan protists. Bottom half displays representatives of metazoans. | |
| Scientific classification | |
| Domain: | Eukaryota |
| Clade: | Amorphea |
| Clade: | Obazoa |
| (unranked): | Opisthokonta |
| (unranked): | Holozoa Lang et al., 2002[1] |
| Clades[4] | |
Incertae sedis | |
| Synonyms | |
| |
Holozoa (from
The holozoan protists play a crucial role in understanding the evolutionary steps leading to the emergence of
Characteristics
Composition
Holozoa is a
- sponges.[9] The mysterious Proterospongia is an example of a colonial choanoflagellate that was thought to be related to the origin of sponges.[10] The affinities of the other single-celled holozoans only began to be recognized in the 1990s.[11]
- pseudopods.[13]
- Syssomonas multiformis. These organisms have varied shapes, including cellular aggregations, amoebae, flagellates, and amoeboflagellates.[6]
- Tunicaraptor unikontum is the newest discovered clade, whose position within Holozoa has yet to be resolved. It is a flagellate with a specialized "mouth" structure absent in other holozoans.[2]
Genetics
The first
Evolution
Phylogeny
Holozoa, along with a clade that contains
Opisthokonta
|
|
||||||||||||||||||||||||||||||||||||||||
| 1250 Mya |

Uncertainty remains around the relationship of the two most basal groups, Ichthyosporea and Pluriformea.[4] They may be sister to each other, forming the putative clade Teretosporea.[19] Alternatively, Ichthyosporea may be the earliest-branching of the two, while Pluriformea is sister to the Filozoa clade comprising filastereans, choanoflagellates and animals. This second outcome is more strongly supported after the discovery of Syssomonas.[2][6]
The position of Tunicaraptor, the newest holozoan member, is still unresolved. Three different phylogenetic positions of Tunicaraptor have been obtained from analyses: as the sister group to Filasterea, as sister to Filozoa, or as the most basal group of all Holozoa.[2][20]
Unicellular ancestry of animals

The quest to elucidate the
- β-catenin).[7]
- ECM-related proteins, involved in the formation of the extracellular matrix, are present in other holozoans (e.g. laminins, collagens and fibronectins).[22]
- A considerable portion of animal transcription factors (TF) is already present in unicellular holozoans, including some TF classes previously thought to be animal-specific (e.g. p53 and T-box).[7]
Additionally, many biological processes seen in animals are already present in their unicellular relatives, such as sexual reproduction and gametogenesis in the choanoflagellate Salpingoeca rosetta and several types of multicellular differentiation.[7]
Fossil record

A
Proposed
Taxonomy
History
Prior to 2002, a relationship between
Holozoa was first recognized as a clade in 2002 through a
Holozoa has since been supported as a robust clade by every posterior analysis,[20] even after the discovery of more taxa nested within it (namely Filasterea since 2008,[13] and the pluriformean species Corallochytrium and Syssomonas since 2014[25] and 2017[6] respectively). As of 2019, the clade is accepted by the International Society of Protistologists, which revises the classification of eukaryotes.[4]
Classification
In classifications that use traditional
- Holozoa Lang et al. 2002
- Incertae sedis: †Bicellum brasieri Strother & Wellman 2021[3]
- Tunicaraptor Tikhonenkov, Mikhailov, Hehenberger, Karpov, Prokina, Esaulov, Belyakova, Mazei, Mylnikov, Aleoshin & Keeling 2020[2]
- Ichthyosporea Cavalier-Smith 1998 [Mesomycetozoea Mendoza et al. 2002]
- Dermocystida Cavalier-Smith 1998
- Ichthyophonida Cavalier-Smith 1998
- Pluriformea Hehenberger et al. 2017
- Corallochytrium Raghu-Kumar 1987
- Syssomonas Tikhonenkov, Hehenberger, Mylnikov & Keeling 2017
- Filozoa Shalchian-Tabrizi et al. 2008
- Filasterea Shalchian-Tabrizi et al. 2008
- Capsaspora Hertel, Bayne & Loker, 2002
- Ministeria Patterson et al. 1993
- Pigoraptor Tikhonenkov et al. 2017
- Txikispora Urrutia, Feist & Bass 2022[12]
- Choanozoa Brunet & King 2017 [Choanozoa Cavalier-Smith et al. 1991 (P)][a]
- ChoanoflagellataKent 1880–1882 [Choanoflagellatea Cavalier-Smith 1997 emend. Cavalier-Smith 1998]
- Craspedida Cavalier-Smith 1997, emend. Nitsche et al. 2011
- Acanthoecida Cavalier-Smith 1997, emend. Nitsche et al. 2011
- MetazoaHaeckel 1874, emend. Adl et al. 2005 [Animalia Linnaeus 1758]
- PoriferaGrant 1836
- Placozoa Grell 1971
- Ctenophora Eschscholtz 1829
- Cnidaria Hatschek 1888
- Bilateria Hatschek 1888
- Filasterea Shalchian-Tabrizi et al. 2008
Notes
References
- ^ PMID 12401173.
- ^ PMID 32976804.
- ^ PMID 33852871.
- ^ PMID 30257078.
- .
- ^ PMID 28648822.
- ^ S2CID 30709486.
- PMID 16151185.
- ISBN 978-3-319-28147-6.
- ISBN 9780429351907.
- PMID 8876236.
- ^ S2CID 240422937.
- ^ PMID 18461162.
- .
- hdl:2027.42/62649.
- PMC 7260361.
- PMID 21810989.
- PMID 26365255.
- PMC 5560861.
- ^ PMID 33622103.
- PMC 7533066.
- PMID 23942320.
- S2CID 4448316.
- S2CID 39813961.
- ^ Torruella G, de Mendoza A, Grau-Bové X, Donachie S, Pérez-Cordón G, Sitjà-Bobadilla A, Paley R, Manohar CS, Nichols K, Eme L, del Campo J (2014). "Phylotranscriptomics reveals ancient and convergent features in Corallochytrium and Ministeria (Holozoa, Opisthokonta)". Phylogeny and evolutionary perspective of Opisthokonta protists (PDF) (PhD thesis). Vol. 75. Universitat de Barcelona. pp. 1–9.
- PMID 34940909.
- PMID 23085100.